
Abstract
Degradation of different cargo by macroautophagy is emerging as a highly selective process which relies upon specific autophagy receptors and adapter molecules that link the cargo with the autophagic molecular machinery. We have recently reported that the large phsophatidylinositol-3-phosphate (PtdIns(3)P)-binding protein Alfy (Autophagy-linked FYVE protein) is required for selective degradation of aggregated proteins. Although depletion of Alfy inhibits Atg5-dependent aggregate degradation, overexpression of Alfy results in Atg5-dependent aggregate clearance and neuroprotection. Alfy-mediated degradation requires the ability of Alfy to directly interact with Atg5. This ability to interact with the core autophagic machinery may cause Alfy to diminish the responsiveness to non-selective autophagic degradation as measured by long-lived protein degradation. Thus, increasing Alfy-mediated protein degradation may be beneficial in some organs, but may be detrimental in others.
Acknowledgements
We would like to acknowledge the following foundations for their generous support: NINDS RO1NS050199, RO1NS063973 and Parkinson's disease foundation (A.Y.); and the Norwegian Council of Research and the Norwegian Cancer Society (A.S.).
Figures and Tables
Figure 1 Modulation of Alfy levels permits starvation-mediated macroautophagy, but modulates the degree of proteolysis in response to starvation. (A) Long-lived protein degradation (LLPD) upon depletion of Alfy by siRNA. HeLa cells were transfected with a scramble siRN A sequence (siCtrl) or a sequence against Alfy (siAlfy). 48 h later, long-lived proteins were labeled with [14C]-valine. Cells were placed in complete media (CM), Hank's Basic Salt Solution (HBSS) with 10 mM HEPES (Starve) or HBSS with 10 mM HEPES plus 10 mM 3-methyladenine (Starve+3MA). Under all conditions 10 mM cold valine was also supplemented. After 4 to 6 h, cells were collected and processed by TCA precipitation and measured for proteolysis as previously described in reference Citation20. ANOVA revealed that upon Alfy depletion there was a significant increase of proteolysis under starvation conditions (Starve, F(1,22) = 5.430; *p = 0.0294). There was no significant difference under nonstarved (CM, F(1,22) = 0.101; p = 0.9782) or starvation + 3MA treated (Starve+3MA, F(1,6) = 1.999; p = 0.2071) conditions (N.S. = ‘not significant’). In both siCtrl and siAlfy transfected cells, starvation led to a significant increase in % proteolysis (siCtrl, p = 0.0207; siAlfy, p = 0.0001) but not in the presence of 3MA (siCtrl, p = 0.8878; siAlfy, p = 0.45). (B) LLPD upon overexpression of Alfy. HeLa cells were transfected with an empty vector (Mock) or a vector encoding the p62- and Atg5-binding C terminus of Alfy (amino acids 2,285–3,526) that was previously shown to increase aggregate-clearance (Alfy C).Citation20 LLPD was monitored as described in (A). ANOVA revealed that overexpression of Alfy leads to significantly less proteolysis upon starvation (Starve, F(1,4) = 8.047; *p = 0.0470). There was no significant difference under unstarved (CM, F(1,4) = 0.115; p = 0.7511) or starvation + 3MA treated (Starve+3MA, F(1,6) = 0.510; p = 0.5145) conditions. In both Mock and Alfy C transfected cells, starvation led to a significant increase in % proteolysis (Mock, p = 0.0001; Alfy C, p = 0.0221) which was significantly inhibited by 3MA (Mock, p = 0.001; Alfy C, p = 0.0473).
![Figure 1 Modulation of Alfy levels permits starvation-mediated macroautophagy, but modulates the degree of proteolysis in response to starvation. (A) Long-lived protein degradation (LLPD) upon depletion of Alfy by siRNA. HeLa cells were transfected with a scramble siRN A sequence (siCtrl) or a sequence against Alfy (siAlfy). 48 h later, long-lived proteins were labeled with [14C]-valine. Cells were placed in complete media (CM), Hank's Basic Salt Solution (HBSS) with 10 mM HEPES (Starve) or HBSS with 10 mM HEPES plus 10 mM 3-methyladenine (Starve+3MA). Under all conditions 10 mM cold valine was also supplemented. After 4 to 6 h, cells were collected and processed by TCA precipitation and measured for proteolysis as previously described in reference Citation20. ANOVA revealed that upon Alfy depletion there was a significant increase of proteolysis under starvation conditions (Starve, F(1,22) = 5.430; *p = 0.0294). There was no significant difference under nonstarved (CM, F(1,22) = 0.101; p = 0.9782) or starvation + 3MA treated (Starve+3MA, F(1,6) = 1.999; p = 0.2071) conditions (N.S. = ‘not significant’). In both siCtrl and siAlfy transfected cells, starvation led to a significant increase in % proteolysis (siCtrl, p = 0.0207; siAlfy, p = 0.0001) but not in the presence of 3MA (siCtrl, p = 0.8878; siAlfy, p = 0.45). (B) LLPD upon overexpression of Alfy. HeLa cells were transfected with an empty vector (Mock) or a vector encoding the p62- and Atg5-binding C terminus of Alfy (amino acids 2,285–3,526) that was previously shown to increase aggregate-clearance (Alfy C).Citation20 LLPD was monitored as described in (A). ANOVA revealed that overexpression of Alfy leads to significantly less proteolysis upon starvation (Starve, F(1,4) = 8.047; *p = 0.0470). There was no significant difference under unstarved (CM, F(1,4) = 0.115; p = 0.7511) or starvation + 3MA treated (Starve+3MA, F(1,6) = 0.510; p = 0.5145) conditions. In both Mock and Alfy C transfected cells, starvation led to a significant increase in % proteolysis (Mock, p = 0.0001; Alfy C, p = 0.0221) which was significantly inhibited by 3MA (Mock, p = 0.001; Alfy C, p = 0.0473).](/cms/asset/874125a2-7185-4467-bdf1-996795ae8fba/kaup_a_10914234_f0001.gif)
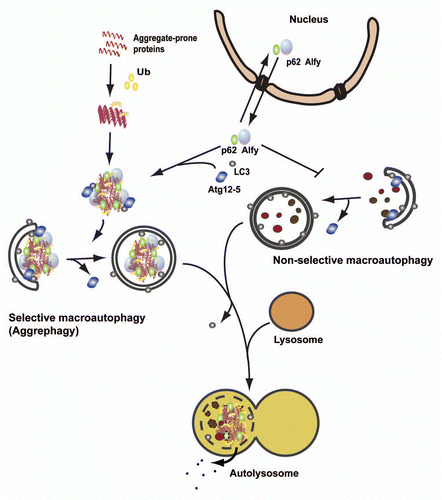
Figure 2 Schematic comparison of selective macroautophagy of aggregate-prone proteins (aggrephagy) and nonselective macroautophagy. Aggregate-prone proteins become ubiquitinated and start to oligomerize. The ubiquitin-binding autophagy receptor p62, together with the large scaffolding protein Alfy, drives the formation of larger aggregates that are targeted for autophagic degradation through interaction of p62 and Alfy with the core autophagic machinery (LC3 and the Atg12-5 complex, respectively). Alfy also binds to PtdIns(3)P in the autophagic membrane and may facilitate binding of Atg12-5 to the membrane-associated Atg16, creating the Atg12-5-Atg16 complex, which might work as an E3-like ligase to permit LC3 conjugation to PE in the membrane, and autophagosomes to form closely around the inclusion. Alfy becomes recruited from the nucleus to cytoplasmic protein aggregates formed upon cellular stress in a p62-dependent manner. Whereas overexpression of Alfy or its p62-, Atg5- and PtdIns(3)P-binding C terminus leads to increased aggrephagy and decreased nonselective starvation-induced macroautophagy, depletion of Alfy has the opposite effect. Thus, nucleocytoplasmic shuttling of Alfy might be a way to regulate the level of aggrephagy versus nonselective macroautophagy.
Addendum to: