Abstract
Bacillus megaterium, the “big beast”, is a Gram-positive bacterium with a size of 4 x 1.5 µm. During the last years, it became more and more popular in the field of biotechnology for its recombinant protein production capacity. For the purpose of intra- as well as extracellular protein synthesis several vectors were constructed and commercialized (MoBiTec GmbH, Germany). On the basis of two compatible vectors, a T7 RNA polymerase driven protein production system was established. Vectors for chromosomal integration enable the direct manipulation of the genome. The vitamin B12 biosynthesis of B. megaterium served as a model for the systematic development of a production strain using these tools. For this purpose, the overexpression of chromosomal and plasmid encoded genes and operons, the synthesis of anti-sense RNA for gene silencing, the removal of inhibitory regulatory elements in combination with the utilization of strong promoters, directed protein design, and the recombinant production of B12 binding proteins to overcome feedback inhibition were successfully employed. For further system biotechnology based optimization strategies the genome sequence will provide a closer look into genomic capacities of B. megaterium. DNA arrays are available. Proteome, fluxome and metabolome analyses are possible. All data can be integrated by using a novel bioinformatics platform. Finally, the size of the “big beast” B. megaterium invites for cell biology research projects. All these features provide a solid basis for challenging biotechnological approaches.
The “Big Beast”
In 1884 de Bary named the Gram-positive bacterium Bacillus megaterium the “big beast” due to its large size.Citation1 With a cell length of up to 4 µm and a diameter of 1.5 µm, B. megaterium belongs to one of the biggest known bacteria. Compared to the model organism for Gram-negative bacteria, Escherichia coli, it has an up to 100-times higher volume (). Due to the dimensions of the vegetative form and spores, B. megaterium is an ideal model system for morphological research. It is well suited for the investigation of cell-wall biosynthesis, cytoplasmic membrane formation and the sporulation process including spore structure and organization.Citation2 In the 1960s, prior to the development of Bacillus subtilis, B. megaterium was “the” model organism of choice for intensive studies on biochemistry, sporulation and bacteriophages of Gram-positive bacteria.Citation3–Citation6 In 1994, Patricia Vary published the review “Prime-time for Bacillus megaterium.”Citation7 In a recent review from 2007, 13 years later, she repeated this sentence and added “… now it is becoming true.”Citation8
First Steps: Bacillus megaterium and Biotechnology
Due to its application for the production of biotechnological relevant substances, B. megaterium is of general interest for this branch of industry.Citation9 Beside its growth on cheap substrates, the non-pathogenic B. megaterium, in contrast to E. coli, does not produce endotoxins associated with an outer membrane.Citation7 Further, it does not possess any obvious alkaline proteases degrading recombinant gene product. Hence, it was sucessfully used for the production of several α- and β-amylases important for starch modification reactions in the baking industry and of penicillin amidase essential for the synthesis of novel β-lactam antibiotics.Citation10 It is also being used industrially for the intracellular production of HIV antigen by the Abbott Laboratories.Citation11
Sweet Life: Xylose Induced Gene Expression
In 1991, Rygus and Hillen pushed B. megaterium research with the introduction of a strong and xylose-inducible promoter found in the genome of B. megaterium. Under the strict control of the repressor protein XylR, the system was used for the controllable production of intracellular recombinant proteins like β-galactosidase, glucose dehydrogenase, formate dehydrogenase and toxin A.Citation12–Citation15 This promoter provided the basis for the effective and commercialized xylose-inducible vector system for B. megaterium: “…one of the most efficient expression systems described in any organism so far” (MoBiTec, Göttingen, Germany).
Express Yourself: Advanced Protein Production using B. megaterium
A novel series of vectors based on the xylose-inducible system was constructed.Citation16 All vectors belonging to this “1622” series have an identical multiple cloning site. This feature allows for simple subcloning in various of these vectors. Upstream or downstream of the MCS the coding sequences of small affinity tags, His6- or StrepII-tag, where introduced. The production of GFP using all vector constructs was successfully tested with a maximal GFP recovery of up to 17.9 mg/L in shaking flask cultures.Citation16 The GFP fusion proteins containing an N- or C-terminal His6- or StrepII-tagCitation17 were easily purified by affinity chromatography. Up to 10.8 mg GFP per liter cell culture were achieved. For upscaling the intracellular production process, GFP was produced under glucose limited high cell density conditions in a 2 liter bioreactor. The induced GFP synthesis, which could be easily pursued via the green fluorescence (), led to 274 mg GFP per liter cell culture. Further, these experiments clearly showed that the xylose-inducible promoter is strictly controlled as no GFP was produced prior induction of gene expression by xylose.Citation16 Another interesting feature was observed: Only 73% of the genetically identical B. megaterium cells showed GFP production while 15% did not (, B and D).Citation16 This culture heterogeneity was described in a different context for B. subtilis before.Citation18
The Big Picture: Cell Biology with B. megaterium
Within the last years, the in vivo observation of cellular processes (“live cell imaging”) in the bacterial cell became more and more common. Much research was done on the Gram-positive organism B. subtilis especially by Peter Graumann's group and others.Citation20–Citation22 For the transfer of these methodologies to the big bug B. megaterium, the intracellular production of recombinant green fluorescent protein (GFP) fusion proteins was a major prerequisite. Due to the size of B. megaterium, a higher resolution of microscopic pictures is expected.
Inside Out: Recombinant Protein Production and Secretion into the Growth Medium
In contrast to the intracellular production of recombinant proteins, the secretion of proteins into the surrounding growth medium provides an elegant and easy method to directly purify heterologous proteins from the medium after cell removal without cell disruption. Next, a series of secretion vectors based on the xylose-inducible vector system () including the sequence of the signal peptide of the lipase A from B. megaterium were constructed using a levansucrase from Lactobacillus reuteri as a model protein.Citation23 The production and secretion of up to 4 mg levansucrase per liter culture medium was observed. Analogously, 1 mg of His-tagged and 0.7 mg of Strep-tagged levansucrase were purified directly from the growth medium.Citation23,Citation24 The efficiency of the established secretion system was further demonstrated by secretion of 6,098 U/l of a hydrolase from Thermobifida fusca,Citation25 the recombinant secretion of 40 mg per liter of penicillin amidase from B. megaterium,Citation26 the export of a keratinase with an activity of 166,200 U/LCitation27 as well as the production and secretion of 400 µg per liter of recombinant single chain Fv antibody fragments.Citation28 All vectors of the secretion series are commercialized by the MoBiTec GmbH (Göttingen, Germany).
In 2004, Nahrstedt et al. described the detection of the signal peptidase SipM in the genome of B. megaterium necessary for the removal of the signal peptide of secreted proteins.Citation29 Recombinant protein secretion of (1,3-1,4)-β-glucanase was increased up to 44% when recombinant SipM was co-produced in B. megaterium. Then, one year later, in 2005, these studies were extended on the secretion of a recombinant 200 kDa protein.Citation19 The dextransucrase DsrS from Leuconostoc mesenteroides secreted into the culture medium of B. megaterium showed an activity of up to 28,600 units per liter. By coproduction of SipM and DsrS, encoded on one vector, the extracellular amount of DsrS was increased 3.7-fold.Citation30
Virus Helps: T7 RNA Polymerase Driven Protein Production in B. megaterium
The B. megaterium gene expression system was completed by the viral T7 RNA polymerase-dependent promoter. This demanded a stable two vector system. However, the origin of replication of all members of the constructed intra- as well as the extracellular vectors was derived from pBC16,Citation31 which is replicated by the rolling circle mechanism.Citation32 For the stable presence of two plasmids in one bacterial cell, origins of replication belonging to different compatibility classes are necessary. B. megaterium strain QM B1551 carries seven different plasmids (pBM100–pBM700).Citation33 One of the corresponding replicons, repBM100,Citation34 was chosen for a second in parallel replicating plasmid in B. megaterium.Citation35 While pBC16 derivatives were found to have 50–100 copies per cell, 135 copies of pBM100 are present.Citation33,Citation36 One vector is carrying the gene encoding the T7 RNA-polymerase under control of the xylose-inducible promoter, while the other vector encodes the target protein under control of the T7 RNA dependent promoter. Using GFP as a model protein, almost 50 mg per liter cell culture were produced.Citation35
Pretty in Pink: Vitamin B12 Production with B. megaterium
Next, we decided to directly optimize the vitamin B12-production with the natural producer B. megaterium.Citation37–Citation39 Different individual manipulation steps were followed. Beside the genetic manipulation of the existing vector DNA, further direct manipulations of the genetic information of B. megaterium were demanded.Citation38 The used strategies illustrate the versatility of this bacterium as a biotechnological toolbox. Recombinant overexpression of single genes (cbiX, sirA, fnr and optimized hemA) increased the vitamin B12 concentration up to 13.5-fold. The integration of the strong xylose-inducible promoter upstream of the hemAXCDBL operon and directed enzyme engineering of HemA to HemAKK by an exchange of two amino acid residues to enhance enzyme stability increased the B12 content up to almost 40-fold (). For this purpose, integration vectors based on the temperature sensitive origin of replication of pE194tsCitation40 were constructed.Citation37,Citation38 The overproduction of the cbiI operon starting from cbiX by chromosomal promoter integration increased the vitamin B12 content almost 20-fold. The biosynthesis of all tetrapyrroles including vitamin B12 and heme uses the same precursor molecule, uroporphyrinogen III. After uroporphyrinogen III, the biosynthetic pathways of vitamin B12 and heme diverge. To repress the flux of this metabolite to the heme biosynthetic path, an antisense RNA strategy to inactivate the hemZ mRNA involved in this competing pathway was established. This was one of the rare examples reported for a successfully employed antisense RNA strategy in bacteria so far. Moreover, overproduction of the recombinant vitamin B12 binding proteins MetH, GlmS and RtpR, respectively, to prevent vitamin B12 dependent feedback inhibition, successfully enhanced the vitamin content almost 5-fold. Here, the twin arginine transport (TAT) system, where folded proteins are secreted, could provide an elegant possibility to export the cofactor vitamin B12 bound to its corresponding vitamin B12 binding domains.Citation41,Citation42
Sometimes More is Less
All directed single manipulations to enhance the cellular vitamin B12 content were successful. The intracellular vitamin B12 amount was increased up to almost 40-fold. Next, the individual strategies were combined. The genetic background of B. megaterium strain HBBm1 containing the xylose-inducible promoter upstream of the hemAKKXCDBL operon (40-fold increase) was combined with the recombinant production of the antisense mRNA of hemZ (1.2-fold increase) and with the recombinant overproduction of sirA (4.5-fold increase), respectively. By simple multiplication, a 48-fold and a 180-fold increase of B12 production, respectively, was expected. But the experiments proved these assumptions wrong. The vitamin B12 content corresponding to the optical density (µg L−1 OD−1578 nm) showed a maximal enhancement of only 33- and 37.2-fold, respectively. And the volumetric amount was even worse. Only maximal values increased 13.6- and 19-fold, respectively, were detected. This was the same phenomenon observed for cells carrying two vectors overproducing GFP. Especially the production of two proteins mediating antibiotic resistance together with the DNA replication of the plasmids seemed to provide stress to the cells.Citation43 Due to this increased metabolic burden for the B. megaterium cell, the cells grew slower, did not reach higher optical densities and further, were limited in their protein production process.
Looking into the Magic Gene Forest
During the last years, the sequences of more and more bacterial genomes were made available. This amount of data provides the possibility to have a systematic look into the genes present as well as in the regulatory areas between them. Currently, the genome sequences of two biotechnological important B. megaterium strains are being obtained (unpublished data). For an easy access to the genomic data the MegaBac platform was set up, accessible under megabac.tu-bs.de.Footnote* Knowledge of the genetic potential of B. megaterium provides the possibility to complete the whole picture of vitamin B12 biosynthesis as well as several other biochemical pathways with impact on the productivity of the host cell. Insights into non-coding regions will give details of regulatory mechanisms. Gene regulating networks can be deduced using the PRODORIC database and the VIRTUAL FOOTPRINT software tool developed in our group (www.prodoric.de).Citation44 Visualization is achieved via ProdoNet.Citation45
The Next Revolution: “Systems Biotechnology”
Now, with help of the genome information, the introduction of modern high-throughput technologies as transcriptomics, proteomics and metabolomics to perform a broad “systems biotechnology” approach is possible. However, in our opinion, “systems biotechnology” is more than just measuring high throughput data: These data have to be combined to establish bioinformatic models for the prediction of cellular processes.
The genome data have already been used to deduce metabolic networks and to model metabolic fluxes. The models have been confronted with the results from metabolome and fluxome analyses.Citation46,Citation47 Consequently now, DNA microarray and proteomic experiments are the experi mental validation for the proposals. Furthermore, integration of regulatory and metabolic networks is desired. The identification of limiting steps in protein production and the deduction of molecular optimization strategies are the major goals in the future.
Figures and Tables
Figure 1 Electron microscope image of Bacillus megaterium (yellow) and Escherichia coli (red) vegetative cells. B. megaterium and E. coli cells were aerobically cultivated separately in LB medium at 37°C. They were grown until reaching the stationary phase. The two cultures were mixed in the ration of 1:1. Aldehyde-fixed bacteria were dehydrated with a graded series of acetone, critical-point-dried with liquid CO2, and sputter-coated with gold. Samples were examined in a field emission scanning electron microscope (FESEM) Zeiss DSM982 Gemini at an acceleration voltage of 5 kV using the Everhart-Thronley SE-detector and the SE-Inlens-detector in a 50:50 ratio. The image was colored subsequently. Magnification ×15,000. Compared to the E. coli volume of 0.5 µm3 (0.5 × 0.5 × 2), B. megaterium has at least up to 100-times higher volume.
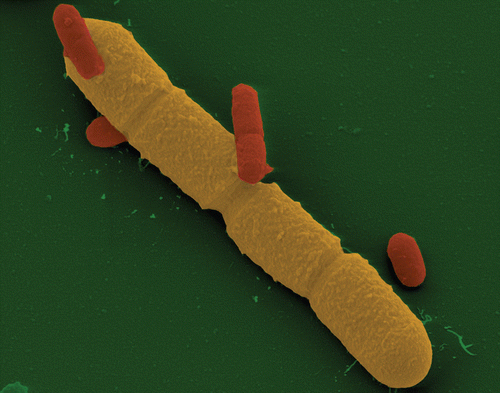
Figure 2 Culture heterogeneity of GFP producing B. megaterium cells (A and B). B. megaterium cells were cultivated on A5 mediumCitation19 agarose pad at 37°C and observed using a Zeiss Axiovert 200 M microscope. Pictures were taken using an AxioCam HR under 630x total magnification (63x objective, 10x ocular). (A) fluorescent image of GFP producing cells; (B) bright image of the same cells overlaid with green-colored fluorescent image. (C) A Biostat B2 bioreactor (B. Braun, Melsungen, Germany) with 2 L working volume connected to an exhaust gas analysis unit (Maihak, Hamburg, Germany) was operated and controlled as described previously.Citation16,Citation19 B. megaterium carrying a plasmid coding for GFP-Strep fusion protein was grown in semi-defined minimal medium at 37°C initially in a batch phase with 4 g/L glucose. At the end of the batch phase an exponential feeding profile was started. GFP was visualized by a lamp emitting blue light and a yellow filter using a digital camera. (D) Results of flow cytometric analysis of bioreactor cultivation. Samples taken from bioreactor cultivation of B. megaterium carrying a plasmid coding for GFP before and 4.6 h after induction of the gfp gene expression were stained with propidium iodide (PI) and analyzed in a FACSCalibur (Benton Dickinson, Belgium): Living cells, no GFP: red; living cells, GFP: green; dead cells, no GFP: black; dead cells, GFP: cyan. Percentages of the subpopulation compared to all cells are given.
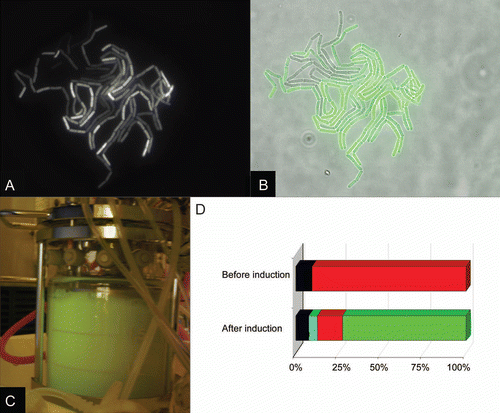
Figure 3 Series of expression plasmids for the (A) extra- and (B) intracellular production of recombinant proteins by B. megaterium. All expression plasmids shown allow parallel cloning of genes of interest into the identical multiple cloning sites (MCS). SP: signal peptide; Tag: Affinity purification tag; CS: protease cleavage site; Term: Terminator; PxylA: promoter of xylA; PT7: T7 RNA polymerase dependent promoter; TEV: tobacco etch virus protease cleavage site; Xa: factor Xa protease cleavage site; SPlipA: signal peptide of the lipase A; SPpac: signal peptide of the penicillin G amidase; TT7: terminator for T7 RNA polymerase.

Figure 4 Shaking flask cultivation of B. megaterium wild type strain and mutant strain HBBm1 (integrated xylose-inducible promoter upstream of the hemAKKXCDBL operon). Wild type bacterial cells and mutant cells were aerobically grown in LB-medium containing 0.23% xylose. Recombinant gene expression was induced with 0.5% xylose at an optical density of 0.4. The picture was taken with a digital camera 5 hours after induction. Left site: wild type cells, right site: Mutant cells. The strong formation of tetrapyrroles is indicated by the red color.

Acknowledgements
We would like to thank Stefan Leupold for fluorescent images. We gratefully acknowledge financial support granted by the Deutsche Forschungsgemeinschaft, especially “Sonderforschungsbereich 578,” and by the “Fonds der Chemischen Industrie.”
Notes
* The platform is still password protected until the publication of the B. megaterium genome sequence. For accessibility requests contact [email protected].
References
- De Bary A. Vergleichende Morphologie und Biologie der Pilze, Mycetozoen und Bacterien 1884; Leipzig Wilhelm Engelmann
- Vary PS. Doi RH, McGloughlin M. Development of genetic engineering in Bacillus megaterium. Biology of Bacilli, Applications to Industry 1992; Boston Butterworth-Heinemann 251 - 310
- Vary PS, Garbe JC, Franzen M, Frampton EW. MP13, a generalized transducing bacteriophage for Bacillus megaterium. J Bacteriol 1982; 149:1112 - 1119
- Vary PS, Halsey WF. Host-range and partial characterization of several new bacteriophages for Bacillus megaterium QM B1551. J Gen Virol 1980; 51:137 - 146
- Lemoigne M, Peaud Lenoel C, Croson M. Assimilation of acetylacetic acid and beta-hydroxybutyric acid by Bacillus megatherium. Ann Inst Pasteur (Paris) 1950; 78:705 - 710
- Clarke NA, Cowles PB. Studies on the host-virus relationship in a lysogenic strain of Bacillus megaterium II. The relationship between growth and bacteriophage production in cultures of Bacillus megaterium. J Bacteriol 1952; 63:177 - 186
- Vary PS. Prime time for Bacillus megaterium. Microbiology 1994; 140:1001 - 1013
- Vary PS, Biedendieck R, Fürch T, Meinhardt F, Rohde M, Deckwer WD, et al. Bacillus megaterium—from simple soil bacterium to industrial protein production host. Appl Microbiol Biotechnol 2007; 76:957 - 967
- Bunk B, Biedendieck R, Jahn D, Vary PS. Flickinger MC. Industrial production by Bacillus megaterium and other Bacilli. Encyclopedia of Industrial Biotechnology 2009; In press
- Panbangred W, Weeradechapon K, Udomvaraphant S, Fujiyama K, Meevootisom V. High expression of the penicillin G acylase gene (pac) from Bacillus megaterium UN1 in its own pac minus mutant. J Appl Microbiol 2000; 89:152 - 157
- Shivakumar AG, Katz L, Cohen LB, Ginsburgh CL, Paul LS, Vanags RI. Expression of heterologous proteins in Bacillus megaterium utilizing sporulation promoters of Bacillus subtilis 1998; USA Abbott Laboratories
- Bäumchen C, Roth AH, Biedendieck R, Malten M, Follmann M, Sahm H, et al. D-mannitol production by resting state whole cell biotransformation of D-fructose by heterologous mannitol and formate dehydrogenase gene expression in Bacillus megaterium. Biotechnol J 2007; 2:1408 - 1416
- Burger S, Tatge H, Hofmann F, Genth H, Just I, Gerhard R. Expression of recombinant Clostridium difficile toxin A using the Bacillus megaterium system. Biochem Biophys Res Commun 2003; 307:584 - 588
- Rygus T, Hillen W. Inducible high-level expression of heterologous genes in Bacillus megaterium using the regulatory elements of the xylose-utilization operon. Appl Microbiol Biotechnol 1991; 35:594 - 599
- Rygus T, Scheler A, Allmansberger R, Hillen W. Molecular cloning, structure, promoters and regulatory elements for transcription of the Bacillus megaterium encoded regulon for xylose utilization. Arch Microbiol 1991; 155:535 - 542
- Biedendieck R, Yang Y, Deckwer WD, Malten M, Jahn D. Plasmid system for the intracellular production and purification of affinity-tagged proteins in Bacillus megaterium. Biotechnol Bioeng 2007; 96:525 - 537
- Lichty JJ, Malecki JL, Agnew HD, Michelson-Horowitz DJ, Tan S. Comparison of affinity tags for protein purification. Protein Expr Purif 2005; 41:98 - 105
- Dubnau D, Losick R. Bistability in bacteria. Mol Microbiol 2006; 61:564 - 572
- Malten M, Hollmann R, Deckwer WD, Jahn D. Production and secretion of recombinant Leuconostoc mesenteroides dextransucrase DsrS in Bacillus megaterium. Biotechnol Bioeng 2005; 89:206 - 218
- Kidane D, Graumann PL. Intracellular protein and DNA dynamics in competent Bacillus subtilis cells. Cell 2005; 122:73 - 84
- Graumann PL. Dynamics of bacterial cytoskeletal elements. Cell Motil Cytoskeleton 2009; 66:909 - 914
- Tadesse S, Graumann PL. Differential and dynamic localization of topoisomerases in Bacillus subtilis. J Bacteriol 2006; 188:3002 - 3011
- Malten M, Biedendieck R, Gamer M, Drews AC, Stammen S, Buchholz K, et al. A Bacillus megaterium plasmid system for the production, export and one-step purification of affinity-tagged heterologous levansucrase from growth medium. Appl Environ Microbiol 2006; 72:1677 - 1679
- Biedendieck R, Beine R, Gamer M, Jordan E, Buchholz K, Seibel J, et al. Export, purification and activities of affinity tagged Lactobacillus reuteri levansucrase produced by Bacillus megaterium. Appl Microbiol Biotech 2007; 74:1062 - 1073
- Yang Y, Malten M, Grote A, Jahn D, Deckwer WD. Codon optimized Thermobifida fusca hydrolase secreted by Bacillus megaterium. Biotechnol Bioeng 2007; 96:780 - 794
- Yang Y, Biedendieck R, Wang W, Gamer M, Malten M, Jahn D, et al. High yield recombinant penicillin G amidase production and export into the growth medium using Bacillus megaterium. Microb Cell Fact 2006; 5:36
- Radha S, Gunasekaran P. Cloning and expression of keratinase gene in Bacillus megaterium and optimization of fermentation conditions for the production of keratinase by recombinant strain. J Appl Microbiol 2007; 103:1301 - 1310
- Jordan E, Hust M, Roth A, Biedendieck R, Schirrmann T, Jahn D, et al. Production of recombinant antibody fragments in Bacillus megaterium. Microb Cell Fact 2007; 6:2
- Nahrstedt H, Wittchen K, Rachman MA, Meinhardt F. Identification and functional characterization of a type I signal peptidase gene of Bacillus megaterium DSM319. Appl Microbiol Biotechnol 2004; 64:243 - 249
- Malten M, Nahrstedt H, Meinhardt F, Jahn D. Coexpression of the type I signal peptidase gene sipM increases recombinant protein production and export in Bacillus megaterium MS941. Biotechnol Bioeng 2005; 91:616 - 621
- Bernhard K, Schrempf H, Goebel W. Bacteriocin and antibiotic resistance plasmids in Bacillus cereus and Bacillus subtilis. J Bacteriol 1978; 133:897 - 903
- Khan SA. Plasmid rolling-circle replication: highlights of two decades of research. Plasmid 2005; 53:126 - 136
- Kieselburg MK, Weickert M, Vary PS. Analysis of resident and transformant plasmids in Bacillus megaterium. Biotechnol 1984; 2:254 - 259
- Kunnimalaiyaan M, Stevenson DM, Zhou Y, Vary PS. Analysis of the replicon region and identification of an rRNA operon on pBM400 of Bacillus megaterium QM B1551. Mol Microbiol 2001; 39:1010 - 1021
- Gamer M, Frode D, Biedendieck R, Stammen S, Jahn D. A T7 RNA polymerase-dependent gene expression system for Bacillus megaterium. Appl Microbiol Biotechnol 2009; 82:1195 - 1203
- Gryczan TJ, Contente S, Dubnau D. Characterization of Staphylococcus aureus plasmids introduced by transformation into Bacillus subtilis. J Bacteriol 1978; 134:318 - 329
- Barg H, Malten M, Jahn M, Jahn D. Barredo JL. Protein and vitamin production in Bacillus megaterium. Microbial Processes and Products 2005; Totowa NJ, USA Humana Press 165 - 184
- Biedendieck R, Malten M, Barg H, Bunk B, Martens JH, Derry E, et al. Metabolic engineering of cobalamin (vitamin B12) production in Bacillus megaterium. Microb Biotechnol 2009; 3:24 - 37
- Martens JH, Barg H, Warren MJ, Jahn D. Microbial production of vitamin B12. Appl Microbiol Biotechnol 2002; 58:275 - 285
- Gruss A, Ehrlich S. Insertion of foreign DNA into plasmids from Gram-positive bacteria induces formation of high-molecular-weight plasmid multimers. J Bacteriol 1988; 170:1183 - 1190
- Tjalsma H, Bolhuis A, Jongbloed JD, Bron S, van Dijl JM. Signal peptide-dependent protein transport in Bacillus subtilis: a genome-based survey of the secretome. Microbiol Mol Biol Rev 2000; 64:515 - 547
- Berks BC, Sargent F, Palmer T. The Tat protein export pathway. Mol Microbiol 2000; 35:260 - 274
- Glick BR. Metabolic load and heterologous gene expression. Biotechnol Adv 1995; 13:247 - 261
- Grote A, Klein J, Retter I, Haddad I, Behling S, Bunk B, et al. PRODORIC (release 2009): a database and tool platform for the analysis of gene regulation in prokaryotes. Nucleic Acids Res 2009; 37:61 - 65
- Klein J, Leupold S, Munch R, Pommerenke C, Johl T, Karst U, et al. ProdoNet: identification and visualization of prokaryotic gene regulatory and metabolic networks. Nucleic Acids Res 2008; 36:460 - 464
- Fürch T, Hollmann R, Wittmann C, Wang W, Deckwer WD. Comparative study on central metabolic fluxes of Bacillus megaterium strains in continuous culture using 13C labelled substrates. Bioprocess Biosyst Eng 2007; 30:47 - 59
- Fürch T, Wittmann C, Wang W, Franco-Lara E, Jahn D, Deckwer W-D. Effect of different carbon sources on central metabolic fluxes and the recombinant production of a hydrolase from Thermobifida fusca in Bacillus megaterium. J Biotechnol 2007; 132:385 - 394