
Abstract
The zebrafish sensory lateral line system has emerged as a powerful model for the mechanistic study of collective cell migration and morphogenesis. Recent work has uncovered the details of a signaling network involving the Wnt/β-catenin, Fgf and Delta-Notch pathways that patterns the migrating lateral line primordium into distinct regions. Cells within these regions exhibit different fundamental behaviors that together orchestrate normal lateral line morphogenesis. In this review, we summarize the signaling network that patterns the migrating lateral line primordium and describe how this patterning coordinates crucial morphogenic cell behaviors.
Figures and Tables
Figure 1 Lateral line embryology visualized using Tg(-8.0cldnb:lyneGFP) embryos. (A) At 24 hpf the primordium (asterisk) has fully migrated onto the somites from its origin near the otic vesicle (OV). (B) The primordium has recently deposited the first proneuromast. (C) Higher magnification of the primordium in (A). Note the rosette shaped proneuromasts forming in the trailing region and the lack of rosettes in the leading region. (D) The post embryonic lateral line is comprised of 4–6 proneuromast in a row along the horizontal myoseptum.
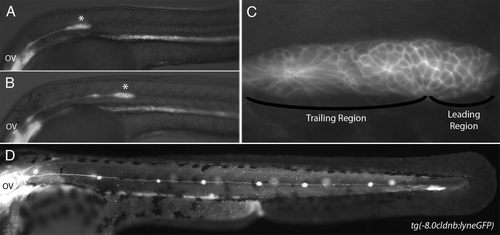
Figure 2 (A) Primordium polarity is maintained by Wnt/β-catenin signaling. The leading region expresses Wnt/β-catenin target genes (red) and the trailing region expresses Fgf target genes (green). Both signaling pathways are active in the central portion of the primordium (yellow). Wnt/β-catenin signaling dependent sef expression represses Fgf signaling in the leading region and Fgf dependent dkk1 expression represses Wnt/β-catenin signaling in the trailing region. Solid lines represent genetic interactions. Dashed lines represent diffusion of secreted factors. (B) Cells in the center of rosette shaped proneuromasts within the primordium express delta and fgf ligands (blue). The remainder of the primordium, with the exception of the leading edge expresses notch receptors (orange). Expression of delta and fgf ligands is restricted to central cells though Delta-Notch mediated lateral inhibition. Cells that constitute the leading edge progenitor pool are outlined in green. Cells in forming proneuromasts are outlined in pink.
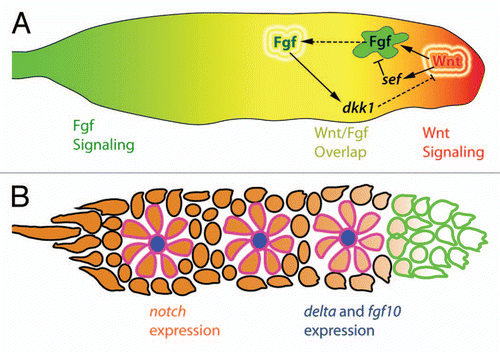
Figure 3 Hypothetical mechanism of apical constriction in forming proneuromasts. Fgf signals emanating from the leading region stimulate the expression of fgf and delta ligands. This expression is restricted to centrally located hair cell progenitors (HC, blue) by Delta-Notch mediated lateral inhibition. Fgf signals from the central hair cell progenitor could potentially interact with Fgf receptors in support cell progenitors (SC, green) leading to the depletion of cortical actin filaments (purple) and radial apical constriction. Apical adherens junction proteins such as ZO-1 and β-catenin (red) accumulate at the center of the forming proneuromast. The forming proneuromast is surrounded by interneuromast cell progenitors (INMC, white) that are outside the range of central cell fgf ligand and therefore do not apically constrict (adapted with permission from Hava et al.Citation32).
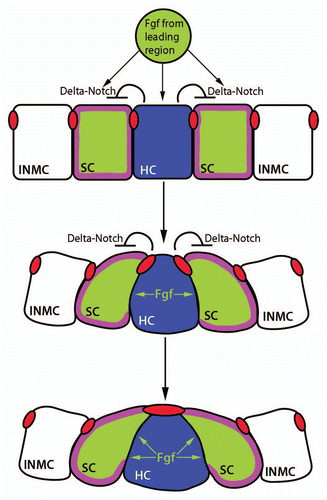
Figure 4 The proliferation-dependent primordium lengthening model of periodic proneuromast deposition. Proliferation in the leading zone (green) displaces cells into the deposition zone (blue) where they begin slowing down and depositing from the primordium. When an entire proneuromast is displaced into the deposition zone it deposits. This leads to a shortened primordium (middle part). Proliferation continues, another proneuromast is displaced into the deposition zone, and the cycle repeats (reproduced with permission from Aman et al.Citation36).
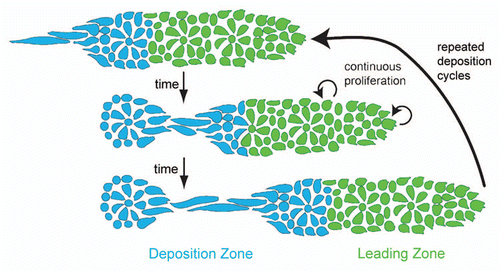
Figure 5 Distribution of classical cadherins in the primordium. e-cadherin (blue) is expressed broadly in the primordium but is downregulated in the leading region and in proneuromast central cells. n-cadherin (orange) is expressed in apically constricted proneuromast cells and in proneuromast central cells. Cells at the primordium periphery and between the proneuromasts within the primordium express e-cadherin but not n-cadherin and represent the presumptive interneuromast cells (adapted with permission from Matsuda and ChitnisCitation20).
