
Abstract
1α,25-dihydroxyvitamin D3 (1,25D) is a powerful differentiation agent, which has potential for treatment of acute myeloid leukemia (AML), but induces severe hypercalcemia at pharmacologically active doses. We have previously shown that carnosic acid (CA), the polyphenolic antioxidant from rosemary plant, markedly potentiates differentiation induced by low concentrations of 1,25D in human AML cell lines. Here, we demonstrated similar enhanced differentiation responses to the 1,25D/CA combination in primary leukemic cells derived from patients with AML, and determined the role of the Nrf2/antioxidant response element (Nrf2/ARE) pathway in these effects using U937 human monoblastic leukemia cells as the model. CA strongly transactivated the ARE-luciferase reporter gene, induced the ARE-responsive genes, NADP(H)-quinone oxidoreductase and the γ-glutamylcysteine synthetase heavy subunit, and elevated cellular glutathione levels. Interestingly, 1,25D potentiated the effects of CA on these activities. Stable transfection of wild-type (wt) Nrf2 resulted in the enhancement, while transfection of dominant-negative (dn) Nrf2 produced suppression of differentiation induced by the 1,25D/CA combination and, surprisingly, by 1,25D alone. These opposite effects were associated with a corresponding increase or decrease in vitamin D receptor and retinoid X receptor-α protein levels, and in vitamin D responsive element transactivation. Cells transfected with wtNrf2 and dnNrf2 also displayed opposing changes in the levels of the AP-1 family proteins (c-Jun and ATF2) and AP-1 transcriptional activity. Pretreatment with AP-1 decoy oligodeoxynucleotide markedly attenuated the differentiation in wtNrf2-transfected cells, suggesting that the pro-differentiation action of Nrf2 is mediated by functional AP-1. Our findings suggest that the Nrf2/ARE pathway plays an important part in the cooperative induction of myeloid leukemia cell differentiation by 1,25D and a plant polyphenol.
Acknowledgements
We thank Dr. Milan Uskokovic, BioXell, Nutley, NJ, for the gift of 1,25α-dihydroxyvitamin D3. This study was partially supported by a NIH grants from the National Cancer Institute RO1-CA-117942-3 to G.P.S. and M.D., RO1-CA-044722-20 to G.P.S. and the Israel Science Foundation grant 778/07 to M.D. and Y.S.
Figures and Tables
Figure 1 Carnosic acid (CA) and 1,25D (D3) cooperate in the induction of differentiation and growth inhibition in U937 cells without inducing cytotoxicity. Cells (4 × 104/ml) were incubated with 0.1% ethanol (vehicle control) or the indicated test agents for 96 h. (A and C) CD11b and CD14 expression was determined by flow cytometry. (B) Wright-Giemsa-stained cytospin smears of the indicated cell samples (magnification, 1,000x). Arrows indicate relatively enlarged cytoplasmic areas of more mature cells with phagosome-like vacuoles and external protrusions. Cell numbers (D) and viability (E) were assessed by the trypan blue exclusion assay. 1,25D at 100 nM was used as a positive control. (A) demonstrates representative bivariate flow cytometric measurements, and (C–E) summarize the data of five independent experiments. *p < 0.05 and $p < 0.001 versus 2.5 nM 1,25D alone. ¶p < 0.01 versus control.
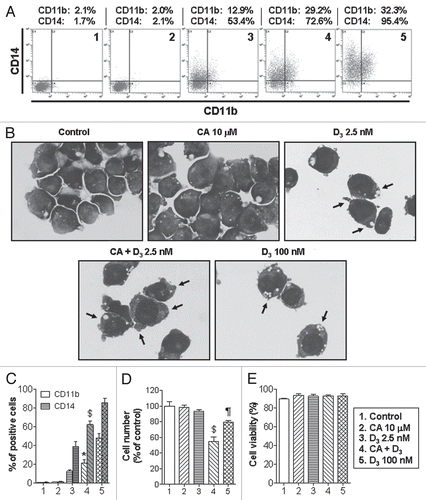
Figure 2 Enhanced differentiation of leukemic blasts from patients with AML following combined treatment with 1,25D and CA ex vivo. Mononuclear cells were isolated from blood specimens of patients with AML-M1 (A), M2 (B) and M4 (C) and incubated at 2 × 105 cell/ml with the indicated agents for 144 h, as described in Materials and Methods. CD11b and CD14 expression was determined by flow cytometry. TPA-stimulated superoxide production was measured in triplicate by the cytochrome c reduction assay (means ± SD).
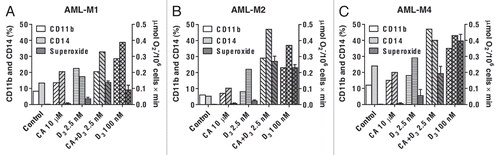
Figure 3 Carnosic acid (CA) and 1,25D (D3) cooperate in the activation of the Nrf2/ARE transcription system and glutathione synthesis in U937 cells. (A) Dose-dependent transactivation of the 4xARE-Luc and γGCSh-ARE4-Luc by CA. Luciferase activity was measured in transiently transfected cells following 24 h incubation with CA, as described in Material and Methods. (B) CA and D3 cooperatively transactivate the 4xARE-Luc and γGCSh-ARE4-Luc, but not the γGCSh-ARE4m-Luc containing mutated ARE4 sequence. Luciferase activity was measured following 24 h incubation with the indicated agents, as in (A). tBHQ was used as the positive control. #p < 0.01 and $p < 0.001 versus CA alone. ‡p < 0.001 versus control. (C) Treatment with CA and D3 does not affect Nrf2 mRNA expression. Cells were incubated with the indicated agents for 1 or 4 h followed by isolation of total mRNA and qRT-PCR analysis, as described in Material and Methods. (D) CA, but not D3, induces elevation of Nrf2 protein levels while the two agents cooperate in the induction of γGCSh. Cells were incubated with the indicated agents for 1 and 6 h (for Nrf2 detection) or 24 h (for γGCSh detection) followed by lysis and western blot analysis, as described in Material and Methods. Calreticulin (CRN) was used as the protein loading control. A representative of three similar blots is shown. (E) CA and D3 cooperatively activate the γGCS mRNA expression. Cells were treated with the indicated agents for 1 and 4 h followed by qRT-PCR, as described above. #p < 0.01 versus CA alone. §p < 0.05 and ¶p < 0.01 versus control. (F) CA and D3 cooperatively elevate the total glutathione levels. Cells were incubated with the indicated agents for 24 h followed by the glutathione reductase recycling assay, as described in the Material and Methods. *p < 0.05 versus CA alone. ¶p < 0.01 and versus control. The treatment groups in (B, C, E and F) are designated “1” to “6” as indicated in (B). (A–C, E and F) The data are the means ± SE of at least three independent experiments performed in duplicate or triplicate.
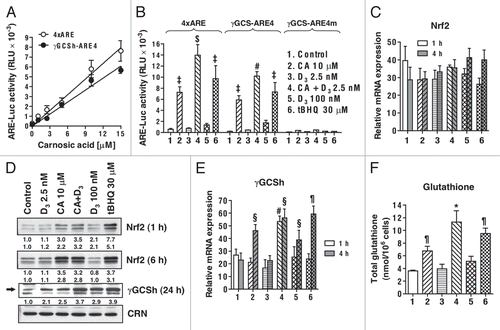
Figure 4 Effects of stable transfection with wtNrf2 or dnNrf2 on the basal and induced levels of Nrf2, NQO1 and γGCSh proteins in U937 cells. (A and B) The indicated wtNrf2, dnNrf2 or empty vector (pEF) transfected clones, mixed pEF clone culture (mix) or untransfected cells (U937) were incubated at 1 × 105 cells/ml with 0.1% ethanol (control) or tBHQ, for 24 h, followed by determination of Nrf2 (A) or NQO1 (B) protein levels by western blotting. (C) The indicated wtNrf2, dnNrf2 or empty vector (pEF) transfected clones or untransfected cells (U937) were incubated, as described above, with ethanol (control) or in the presence of CA or tBHQ, for 24 h, followed by determination of γ-GCS protein levels by western blotting. Calreticulin (CRN) was used as a protein loading control. Representative blots of three similar experiments are shown.
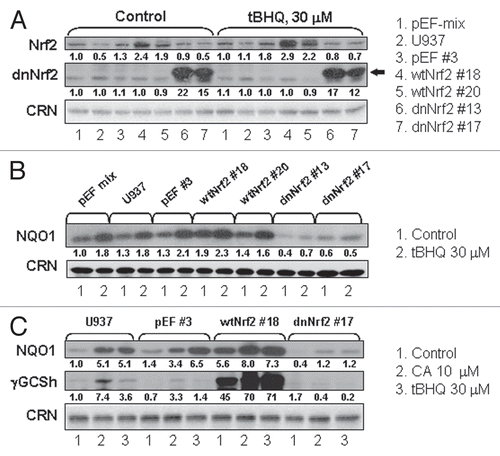
Figure 5 Stable expression of wtNrf2 or dnNrf2 modulates differentiation responses of U937 cells to the 1,25D/CA combination and 1,25D alone. The indicated wtNrf2-, dnNrf2- and empty vector (pEF)-transfected U937 clones and untransfected (U937) cells were incubated with 0.1% ethanol (control), CA, 1,25D (D3) or D3/CA combination for 96 h followed by analysis of CD11b (A) and CD14 (B) expression. Data are the means ± SE of five independent experiments. §p < 0.05; ¶p < 0.01 and ‡p < 0.001 versus corresponding responses of the p-EF#3 cells.
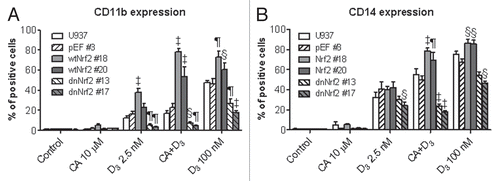
Figure 6 Stable expression of wtNrf2 or dnNrf2 modulates c-Jun, c-Fos and ATF2 protein levels and AP-1 transactivation in U937 cells. The indicated clones of U937 cells (1 × 105 cells/ml) stably transfected with empty vector (pEF#3; A and B), wtNrf2 (A) or dnNrf2 (B) were treated with vehicle (control) or the agents indicated in (C), for 24 h. Whole cell lysates were analyzed by western blotting. Calreticulin (CRN) was used as a protein loading control. Representative blots of three similar experiments are shown. The identification numbers of wtNrf2 and dnNrf2 clones are indicated next to the corresponding blots. (C) TRE × 3-Luc reporter activity was determined in untransfected U937 cells (U937) and in the indicated stable clones following transient transfection with TREx3-Luc and Renilla luciferase and treatment with vehicle (control) or indicated test agent for 24 h. The relative TRE-Luc activity (means ± SE) was calculated from the data of three individual experiments performed in triplicate. The treatment groups in (A and B) are designated “1” to “5” as indicated in (C). §p < 0.05; ¶p < 0.01 and ‡p < 0.001 versus corresponding responses of the pEF#3 cells.
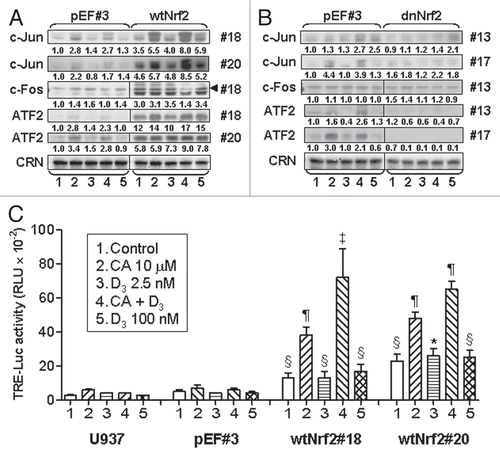
Figure 7 Treatment with AP-1 decoy oligonucleotide blocks the differentiation responses to the 1,25D/CA combination, most robustly in wtNrf2-transfected cells. U937 cells stably expressing empty vector (pEF) or wtNrf2 were preincubated with 7.5 µM TRE-ODN or the mutant mTRE-ODN for 24 h followed by treatment with the indicated agents for an additional 60 h. The expression of CD11b and CD14 was then measured by the bivariate flow cytometric analysis. (A) Comparative read-outs of CD11b and CD14 cell surface expression obtained from ODN-treated pEF #3 and wtNrf2 #18 cells. (B) Averaged CD11b and CD14 expression data from ODN-treated pEF #3 and pEF-mix cells (pEF-CD11b and pEF-CD14, respectively) and from wtNrf2 #18 and wtNrf2 #20 cells (wtNrf2-CD11b and wtNrf2-CD14, respectively). Data are the means ± SE of three independent experiments *p < 0.05 and #p < 0.01 versus response to mTRE-ODN in the same treatment group.
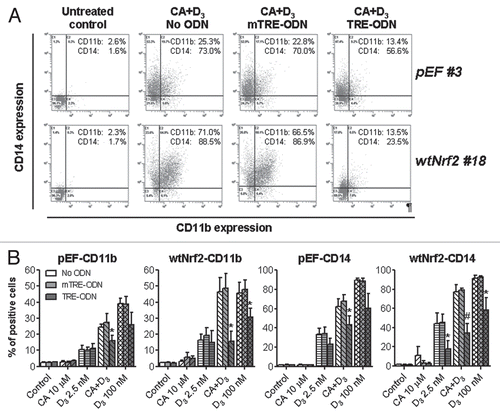
Figure 8 Modulation of VDR and RXRα protein levels and VDRE transactivation by stable expression of wtNrf2 or dnNrf2. The indicated clones of U937 cells (1 × 105 cells/ml) stably transfected with empty vector (pEF#3; A and B), wtNrf2 (A) or dnNrf2 (B) were treated with vehicle (control) or the agents indicated in (C), for 24 h. Whole cell lysates were analyzed by western blotting. Calreticulin (CRN) was used as a protein loading control. Representative blots of three similar experiments are shown. The identification numbers of wtNrf2 and dnNrf2 clones are indicated next to the corresponding blots. Note that the exposure of the RXRα blot from dnNrf2 #13 cells (B) to X-ray film was longer than in the rest of the RXRα blots (A and B) in order to allow visualization of lower RXRα expression in this dnNrf2 clone compared to that in the pEF #3 clone. (C) VDRE-Luc reporter activity was determined in untransfected U937 cells (U937) and in the indicated stable clones following transient transfection with VDREx6-Luc and Renilla luciferase and treatment with vehicle (control) or indicated test agent for 24 h. The relative VDRE-Luc activity (means ± SE) was calculated from the data of three individual experiments performed in triplicate. The treatment groups in (A and B) are designated “1” to “5” as indicated in (C). §p < 0.05; ¶p < 0.01 and ‡p < 0.001 versus corresponding values of the pEF#3 cells.
Figure 9 Suggested modes of the involvement of Nrf2/ARE in the regulation of AML cell differentiation. Plant antioxidants, e.g., CA activate Nrf2 by releasing it from the inhibitory partner, Keap-1. As a result, Nrf2 binds to ARE sequences and heterodimerizes with small Maf proteins leading to the induction of phase II enzymes and other redox regulators, e.g., γGCS, responsible for glutathione synthesis, and thioredoxin. 1,25D can further activate the Nrf2/ARE pathway, possibly by post-transcriptionally activating Nrf2 via phosphorylation by signaling protein kinases (PKC, MAPKs and/or PI-3-K). In addition, 1,25D can probably further enhance γGCS expression by Nrf2/ARE-independent mechanism. The resulting generation of reducing conditions can increase the activity of AP-1 by maintaining the cysteine residues (C in KCR) in their reduced (SH) form, which is required for DNA binding. Ref-1 is a redox regulator.Citation54 KCR is a tripeptide in the DNA binding region of both Jun and Fos. Our data indicate that Nrf2 can also positively regulate the expression AP-1 family proteins (cJun, cFos and ATF-2), further upregulating the AP-1 functional activity. Activated AP-1 can increase the expression of VDR and, perhaps, RXRα, resulting in the enhancement of the transcriptional activity of VDR/RXRα complex. A possibility exists that Nrf2 may also facilitate VDR and RXRα expression in an AP-1-inependent manner. Eventually, the enhancement of VDR function by 1,25D/CA is likely to increase the sensitivity of AML cells to the differentiation-inducing effects of lower concentrations of 1,25D.
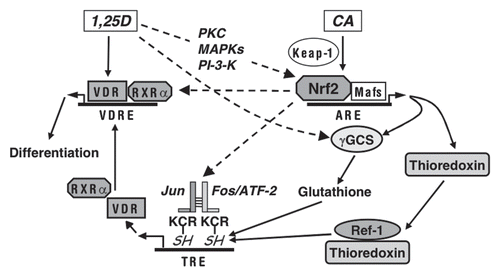
Table 1 Primer sequences used for quantitative RT-PCR