Abstract
Genetic screens in the fruit fly Drosophila melanogaster have identified a class of neoplastic tumor suppressor genes (endocytic nTSGs), which encode proteins that localize to endosomes and facilitate the trafficking of membrane-bound receptors and adhesion molecules into the degradative lysosome. Loss of endocytic nTSGs transforms imaginal disc epithelia into highly proliferative, invasive tissues that fail to differentiate and display defects in cellular apicobasal polarity, adhesion and tissue architecture. As vertebrate homologs of some Drosophila nTSGs are linked to tumor formation, identifying molecular changes in signaling associated with nTSG loss could inform understanding of neoplastic transformation in vertebrates. Here we show that mutations in genes that act at multiple steps of the endolysosomal pathway lead to autonomous activation of the Sav/Wts/Hpo (SWH) transcriptional effector Yki (YAP/TAZ in vertebrates) and the Jun N-terminal kinase (JNK), which is known to promote Yki activity in cells with disrupted polarity. Yki and JNK activity are elevated by mutations at multiple steps in the endolysosomal pathway including mutations in the AP-2σ gene, which encodes a component of the AP-2 adaptor complex that recruits cargoes into clathrin-coated pits for subsequent internalization. Moreover, reduction of JNK activity can decrease elevated Yki-signaling caused by altered endocytosis. These studies reveal a broad requirement for components of the endocytic pathway in regulating SWH and JNK outputs, and place Drosophila endocytic nTSGs into a network that involving two major signaling pathways implicated in oncogenesis.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank M. Gilbert, D. Bilder, G. Halder, M. Scott, R. Fehon, U. Tepass and E. Knust for gifts of fly stocks and antibodies. We also thank the Bloomington Drosophila Stock Center and the Developmental Studies Hybridoma Bank for fly stocks and antibodies. We are grateful to R. Jones and members of the Moberg laboratory for helpful discussions. This work was supported by NIH R01CA123368 to K.H.M. and the Emory BCDB and MTSP Training Grants (T32 GM008367 and T32 GM008169) to B.S.R.
Figures and Tables
Figure 1 ept loss elevates Yki activity. α-β-gal staining to detect the ex-lacZ reporter (red in A; blue in B) in (A) ept or (B) ept,H99 mosaic imaginal eye discs (clones are marked by the absence of GFP).
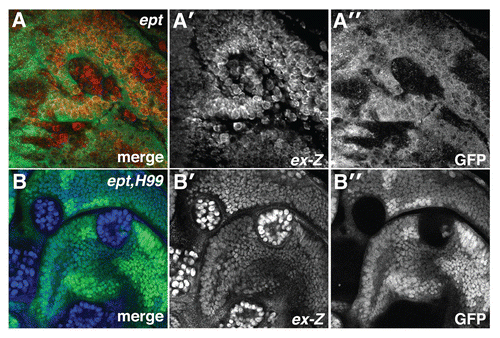
Figure 2 Multiple blocks in the endolysosomal pathway elevate Yki activity. α-β-gal staining of larval imaginal wing discs in which the ex-lacZ SWH-reporter has been placed in the background of (A) en, (B) en > AP-2σ-IR (C) en > Rab5DN (D) en > syx7-IR and (E) en > vps25-IR animals. Tangential and lateral sections are shown for each genotype. Lateral sections were obtained from the dorsal-ventral margin of the wing epithelium as indicated by the line in (A). Posterior compartment is to the right of the dotted line.
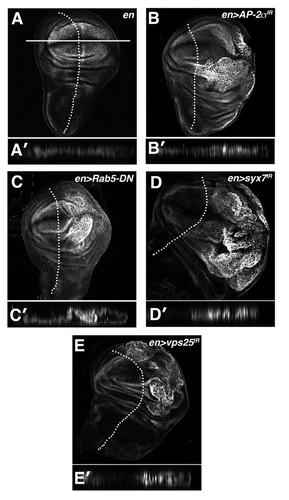
Figure 3 Endolysosomal growth phenotypes are sensitive to the genetic dose of Yki. Light microscopic images of (A) eyFLP;FRT80B/M(3), (A′) eyFLP;Act > CD2 > Gal4, UAS-sav;FRT80B/M(3) and (A″) eyFLP,Act > CD2 > Gal4,UAS-sav;avlCitation1/M(3) adult eyes, in which sav is overexpressed using the Act > CD2 > Gal4 ‘Flp-out technique’. (B and C) Quantitative analysis showing the percent of pupae at 6 d AED (after egg deposition) in the indicated genotypes. A minimum of 50 animals was counted per genotype. (D) α-β-gal staining to detect ex-lacZ reporter in the background of the indicated genotypes: (D) en > yki-IR (D′) en > AP-2σ-IR and (D″) en > AP-2σ-IR,yki-IR. Posterior compartment is to the right of the dotted line.
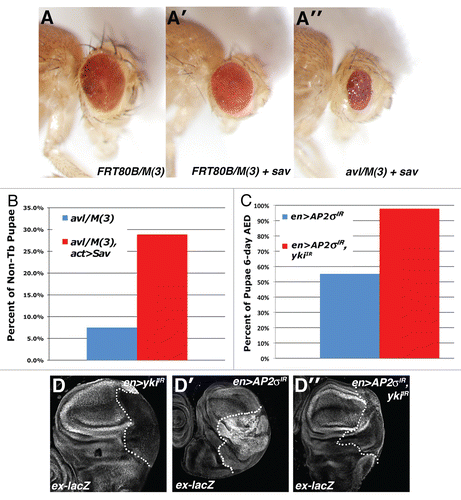
Figure 4 Loss of ept elevates JNK activity. Confocal images of (A–B) eptCitation2 and (C–D) eptx1,H99 clones marked by the absence of GFP stained for (A and C) MMP1 and (B and D) phosphorylated-JNK (p-JNK). Arrows denote p-JNK in mutant clones.
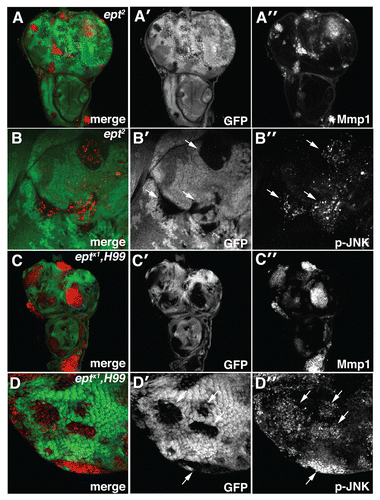
Figure 5 Depletion of AP-2σ, Syx7 or Vps25 elevates JNK activity. Confocal images of larval imaginal wing discs from (A and G) en > AP-2σ-IR, (B and H) en > syx7-IR and (I) en > vps25-IR flies stained for (A–C) MMP1 and (G–I) p-JNK. (D–F) α-β-Gal staining of larval imaginal wing discs in which the puc-lacZ JNK-pathway reporter has been placed into the background of (D) control en >, (E) en > AP-2σ-IR or (F) en > syx7-IR flies. Wing discs are oriented with the posterior compartment to the right.
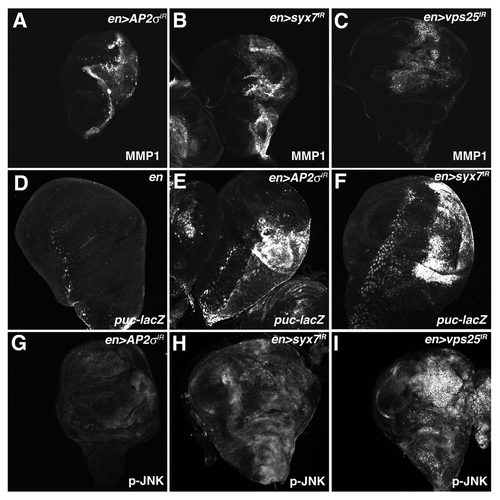
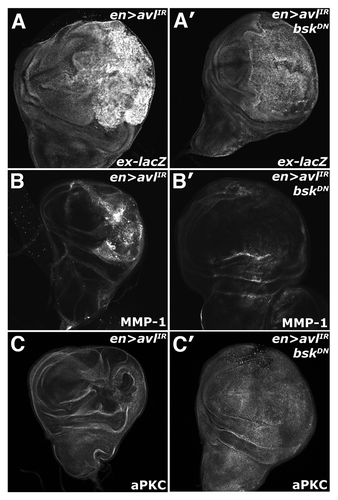
Figure 6 Endolysosomal activation of Yki requires Bsk activity. Confocal images of larval imaginal wing discs in which the ex-lacZ reporter has been placed in the background and of (A–C) en > syx7-IR and (A′–C′) en > syx7-IR, DN-Bsk animals and stained for (A) α-β-gal, (B) MMP1 and (C) aPKC to show disc morphology. Wing discs are oriented with the posterior compartment to the right.