Abstract
Rapamycin is an inhibitor of the mammalian Target of Rapamycin, mTOR, a nutrient-sensing signaling kinase and a key regulator of cell growth and proliferation. While rapamycin and related compounds have anti-tumor activity, a prevalent characteristic of cancer cells is resistance to their anti-proliferative effects. Our studies on nutrient regulation of fetal development showed that hepatocyte proliferation in the late gestation fetal rat is resistant to rapamycin. Extension of these studies to other tissues in the fetal and neonatal rat indicated that rapamycin resistance is a characteristic of normal cell proliferation in the growing organism. In hepatic cells, ribosomal biogenesis and cap-dependent protein translation were found to be relatively insensitive to the drug even though mTOR signaling was highly sensitive. Cell cycle progression was also resistant at the level of cyclin E-dependent kinase activity. Studies on the effect of rapamycin on gene expression in vitro and in vivo demonstrated that mTOR-mediated regulation of gene expression is independent of effects on cell proliferation and cannot be accounted for by functional regulation of identifiable transcription factors. Genes involved in cell metabolism were overrepresented among rapamycin-sensitive genes. We conclude that normal cellular proliferation in the context of a developing organism can be independent of mTOR signaling, that cyclin E-containing complexes are a critical locus for rapamycin sensitivity, and that mTOR functions as a modulator of metabolic gene expression in cells that are resistant to the anti-proliferative effects of the drug.
Acknowledgements
We thank the members of our laboratory who contributed to this work, including Anand Padmanabhan and Rosa Jimenez. These studies were supported by NIH Grants R01 HD24455 and R01 HD35831 (to P.A.G.), and P20 RR017695 (to J.A.S.).
Figures and Tables
Figure 1 The effect of rapamycin on hepatocyte proliferation and mTOR signaling in late gestation fetal and regenerating adult liver. Fetal rats were administered rapamycin (5 µg per fetus) or DMSO vehicle (control) in situ by intraperitoneal injection. The fetuses were replaced, the laparotomy incision was closed and gestation was allowed to continue for an additional 24 h. Adult rats underwent partial hepatectomy following intraperitoneal administration of rapamycin (2.5 mg per kg body weight) or DMSO vehicle. Animals were sacrificed 24 h later. In both cases, BrdU (50 mg per kg body weight) was administered at the time of rapamycin or DMSO administration. (A) The photomicrographs show BrdU immunohistochemical staining of liver sections from representative fetal or adult animals (from ref. Citation25; Boylan et al. J Biol Chem 2001; 276:44457). (B) Livers were analyzed by immunoblotting for phosphorylated (P-S6235/236) and total ribosomal protein S6.
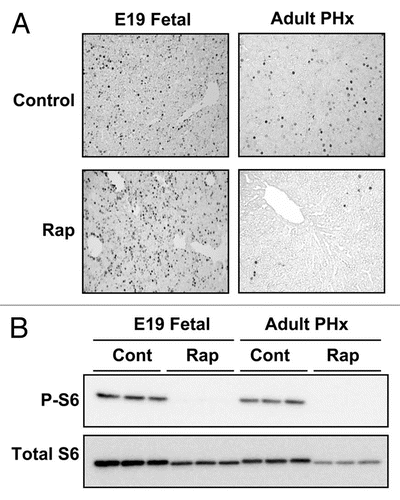
Figure 2 Effect of rapamycin on DNA synthesis in E19 fetal rats. E19 fetuses were injected in situ with DMSO vehicle or rapamycin plus BrdU. The fetuses were replaced and gestation was allowed to continue for 24 h. (A) Fetal whole mounts were fixed in formalin, paraffin-embedded and stained for BrdU. (B) Formalin-fixed, paraffin embedded tissues were stained for P-S6235/236 and counterstained with hematoxylin (from ref. Citation29; Sanders et al. Am J Physiol Cell Physiol 2008; 295:406).
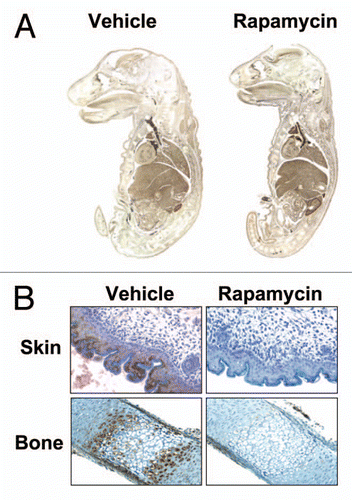
Figure 3 Rapamycin effect on mTOR signaling and cell cycle control in hepatic cell lines. (A) Duplicate samples from five cell lines exposed to rapamycin (50 nM for 24 h) or control vehicle (DMSO) were analyzed by immunoblotting for phosphorylated (P-S6235/236) and total ribosomal protein S6. (B) Similar samples, prepared from H4IIE cells, were analyzed for cyclin E-dependent kinase activity and composition of cyclin E-containing complexes using immunoprecipitation and immunoblotting. Similar studies carried out on four additional cell lines were analyzed for cyclin E-dependent kinase activity (C) and cyclin E-associated p27Kip1 (D). Filled bars represent control vehicle and unfilled bars rapamycin treated cells. Graphs show triplicate analyses as the mean plus 1SD. *p < 0.05 versus corresponding control (from ref. Citation33; Jimenez et al. PLoS ONE 4:7373, 2009).
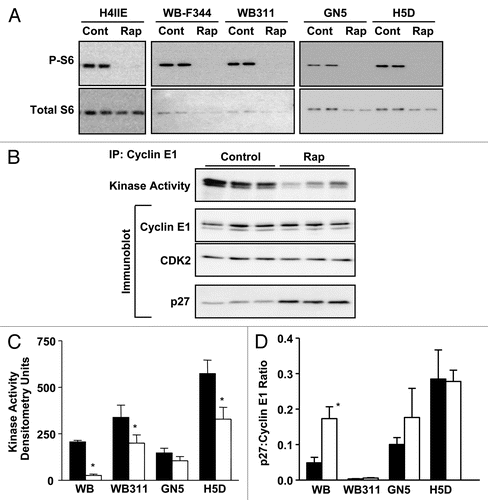
Figure 4 Effect of rapamycin on cyclin E-dependent kinase activity and composition of cyclin E-containing complexes in fetal and adult liver. Animal studies were carried out as for . (A) Triplicate liver samples were analyzed for cyclin E-dependent kinase activity. Filled bars represent DMSO vehicle-injected animals and unfilled bars represent rapamycin-injected animals. Graphs show triplicate analyses as the mean plus 1SD. *p < 0.05 versus corresponding control. (B) Duplicate samples were analyzed for composition of cyclin E-containing complexes.
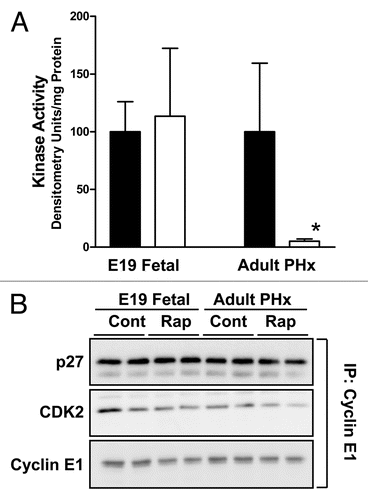
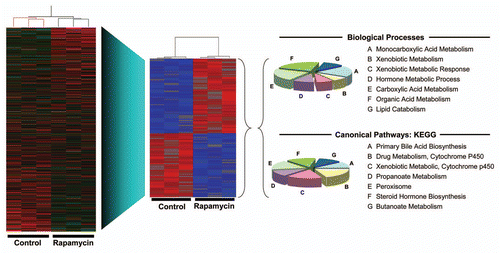
Figure 5 Effect of rapamycin on gene expression in late gestation fetal rat liver. Fetuses were administered rapamycin or DMSO vehicle in situ as described for . Total RNA was prepared from triplicate control and experimental fetuses and analyzed by microarray (Affymetrix® Rat Gene 1.0 ST Array). To the left is shown a heat map and hierarchical clustering for all genes (red, high expression; green, low expression). From this dataset, a subset of genes was identified as showing a significant (p < 0.05) change in expression of at least 1.5-fold in response to rapamycin (red, high expression; blue, low expression). These genes were analyzed by GSEA for biological processes (top) and canonical pathways (bottom). The pie charts show the distribution of genes among categories for which the level of significance was p < 0.005 (40 genes for biological processes and 36 for canonical pathways).