Abstract
We report that simultaneous expression of Akt and angiopoietin-1 (Ang-1) transgenes supported mitogenesis in stem cells with a critical role for microRNA-143 (miR-143) downstream of FoxO1 transcription factor. Mesenchymal stem cells (MSC) from young male rats were transduced with Ad-vectors encoding for Akt (AktMSC) and Ang-1 (Ang-1MSC) transgenes for their individual or simultaneous overexpression (AAMSC; > 5-fold gene level and > 4-fold Akt and Ang-1 protein expression in AAMSC vs. Ad-Empty transduced MSC; EmpMSC). AAMSC had higher phosphorylation of FoxO1, which activated Erk5, a distinct mitogen-induced MAPK that drove transcriptional activation of cyclin D1 and Cdk4. Flow cytometry showed > 10% higher S-phase cell population that was confirmed by BrdU assay (15%) and immunohistology for Ki67 (11%) in AAMSC using EmpMSC as controls. miR array supported by real-time PCR showed induction of miR-143 in AAMSC (4.73-fold vs.. EmpMSC). Luciferase assay indicated a dependent relationship between miR-143 and Erk5 in AAMSC. FoxO1-specific siRNA upregulated miR-143, whereas inhibition of miR-143 did not change FoxO1 activation. However, miR-143 inhibition repressed phosphorylation of Erk5 and abrogated cyclin D1 with concomitant reduction in cells entering cell cycle. During in vivo studies, male GFP+ AAMSC transplanted into wild-type female infarcted rat hearts showed significantly higher number of Ki67 expressing cells (p < 0.05 vs. EmpMSC) 7 days after engraftment (n = 4 animals/group). In conclusion, co-overexpression of Akt and Ang-1 in MSC activated cell cycle progression by upregulation of miR-143 and stimulation of FoxO1 and Erk5 signaling.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Funding
This work was supported by National Institutes of Health (NIH) Grants # R37HL074272; HL-080686; HL-087246 (M.A.) and HL-087288; HL-089535; HL106190-01 (Kh.H.H.).
Figures and Tables
Figure 1 In vitro characterization of MSC for transgenes expression. (A) Flow cytometric analysis of hematopoietic and mesenchymal specific membrane markers CD45, CD29 and CD90 in the purified MSC culture used during the present studies. (B) Double fluorescence immunostaining for Akt (green fluorescence) and Ang-1 (red fluorescence) expression in AAMSC. (C) RT-PCR for Akt and Ang-1 transgene expression in various treatment groups of cells. Densitometry showed significantly higher Akt and Ang-1 in AktMSC, Ang-1MSC and AAMSC as compared with EmpMSC at 48 h after transduction. Transgene expression was normalized to actin. (D) Western blots showing total Akt, pAkt and Ang-1 protein expression standardized to actin in various treatment groups of MSC. Highest level of Akt, pAkt and Ang-1 was observed in AAMSC as compared with Ang-1MSC, AktMSC and EmpMSC.
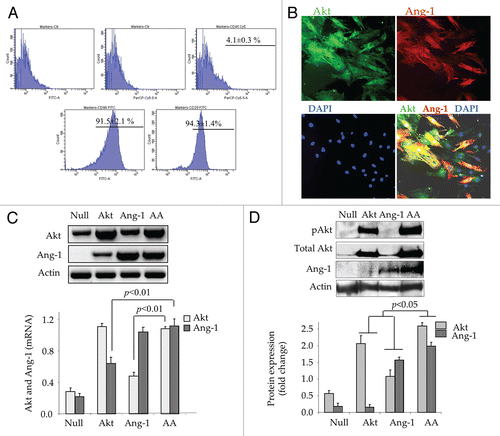
Figure 2 Phosphorylation of FoxO1 and its nuclear exportation in AAMSC. (A and B) Western blots showing significantly higher phosphorylation of FoxO1 at Ser256 with concomitant reduction in total FoxO1 in the whole cell lysate using actin for normalization during densitometry. (C) Representative western blots of FoxO1 and phosphorylated FoxO1 at Ser256 and Thr24 in the nuclear and cytoplasmic fractions of various groups of cells. Whereas Thr24-FoxO1 was mainly activated in cytoplasmic fraction and Ser256-FoxO1 was mainly activated in the nuclear fraction, actin was used as loading control for cytoplasmic fraction and CREB transcription factor was used for loading control in nuclear fraction. (D) Representative images of immunostained AAMSC showing cytoplasmic localization of Thr24-FoxO1 (red fluorescence). No Thr24-FoxO1 was observed in EmpMSC. Nuclei were visualized by DAPI staining (blue). (E and F) Proportion of pErk5 normalized to total Erk5 and total cyclin D1 normalized to actin in the whole cell lysate samples from after respective treatment. Highest level of pErk5 and Cyckin D1 were observed in AAMSC. (G) Representative images of AAMSC immunostained for cyclin D1 expression. Extensive nuclear localization of cyclin D1 (ref fluorescence) was observed in AAMSC. (H) Western blots showing significantly higher expression of Cyclin dependent kinase-4 expression in AAMSC as compared with other treatment groups of cells. Actin was used as an internal control.
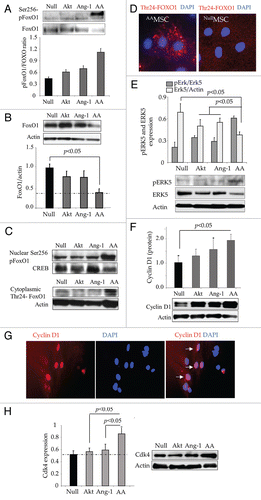
Figure 3 miR-143 expression in AAMSC regulated cell cycle signaling. (A) Heat map showing miR expression profiles in different treatment groups of MSC. (B) Validation of microarray data by Real-time PCR which confirmed significantly higher expression of miR-143 in AAMSC as compared with AktMSC, Ang-1MSC and EmpMSC as controls. (C) Expression of miR-143 was significantly increased in AAMSC. Similarly, when native MSC (without any viral vector transduction) were treated with siFoxO1, miR-143 expression was significantly increased (similar to AAMSC) whereas treatment of native MSC with scramble did not alter miR-143 expression, thus suggesting that miR-143 expression was FoxO1 dependent. (D) Western blots showing FoxO1 was abrogated by co-expression of Akt/Ang-1 in AAMSC as compared with EmpMSC. However, prior treatment of AAMSC with miR-143 specific antagomir did not rescue FoxO1 expression in AAMSC. (E) Western blots showing ectopic expression of miR-143 in native MSC led to abrogation of total Erk5 expression. However, we observed concomitant increase in phosphorylation of Erk5 in comparison with scramble transfected cells. (F) Western blots showing significantly higher expression of cyclin D1 in native MSC with ectopic expression of miR-143. Scramble transfection in AAMSC did not alter cyclin D1 expression. (G and H) Western blots showing significantly higher induction of Erk5 and abrogation of cyclin D1 in native MSC treated with miR-143 specific antagomir as compared with the scramble transfected native MSC as controls (p < 0.05). Although Erk5 was significantly increased after pretreatment with miR-143 specific antagomir, phosphorylation of Erk5 was significantly decreased in AAMSC as compared with scrambled treated AAMSC.
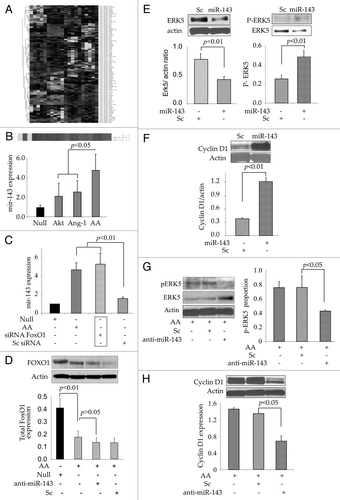
Figure 4 Erk5 is a direct target of miR-143. (A) Computational analysis predicting possible target site of miR-143 conserved in the 3′UTR of Erk5 mRNA. (B) The construction of pEZX-Luc-Erk5 3′UTR luciferase reporter plasmid and precursor miR-143 expression clone. Co-transfection of MSC with pEZX-Luc vector containing Erk5 3′ UTR together with a plasmid encoding miR-143 showed decreased luciferase activity (p < 0.01 vs. pEZX-miR-SC transfected cells). The ratio of luciferase activity was calculated either in the presence or absence of miR-143. (C) Luciferase assay measurement in AAMSC transfected with vectors carrying Luciferase and Erk5 genes.
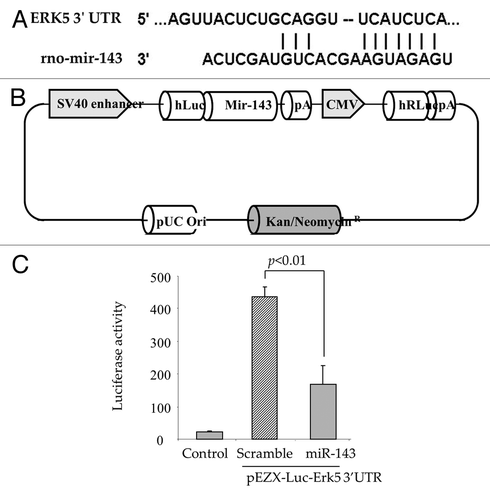
Figure 5 Co-overexpression of Akt/Ang-1 transgenes enhanced cell proliferation. (A) Graph showing significantly higher BrdU uptake in AAMSC as compared with EmpMSC. However, prior treatment of AAMSC with miR-143 antagomir significantly reduced BrdU positivity in AAMSC as compared with scramble (Sc) treated AAMSC. (B) Representative merged fluorescence images of AAMSC immunostained for BrdU uptake (green fluorescence). The nuclei were visualized by DAPI staining (blue fluorescence) and showed nuclear localization of BrdU signals (green fluorescence). White boxed areas in AAMSC and EmpMSC images were magnified for clarity (original magnification = 40x). (C and D) Graph showing significantly higher percentage of AAMSC in G-S phase transition. The cells were stained with propidium iodide and analyzed by FACS at 488 nm. The native (non-transduced) MSC and EmpMSC were used as controls. Prior treatment of the AAMSC with miR-143 antagomir significantly reduced G-S phase transition of the cells. (D) Typical representative histograms from FACS analysis for G-S phase transition of native, AAMSC and AAMSC with miR-143 antagomir treatment or Sc treatment. (E and F) Fluorescence immunostaining of cells from various treatment groups for Ki67 expression (green fluorescence). The nuclei were visualized by DAPI staining which helped to determine nuclear specificity of Ki67 expression (original magnification = 40x). The percentage of Ki67+ cells was significantly increased in AAMSC which was abolished by pretreatment of the cells with miR-143 antagomir. Pretreatment with scramble (Sc) did not alter Ki67 positivity in AAMSC.
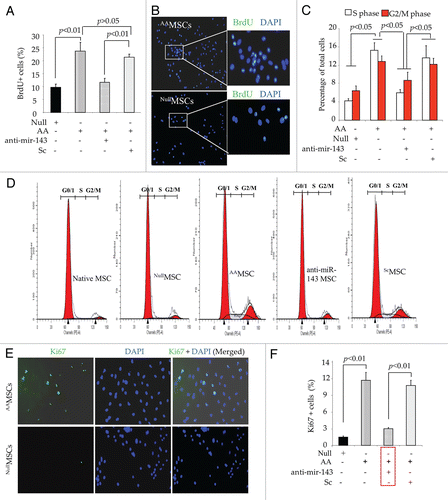
Figure 6 Proliferation of AAMSC post-transplantation in the infarct myocardium. (A) Quantification of Ki67+ positivity in the infarcted myocardium on day 7 after transplantation. The total number of Ki67+ cells was significantly higher in AAMSC transplanted animal hearts as compared with the other treatment groups. (B–D) Representative images of histological sections on day 7 after treatment with (B) EmpMSC and (C) AAMSC. The histological sections were immunostained for Ki67 (red) and GFP (green) antigens. (D) Graph showing higher number of Ki67+/GFP+ cells in AAMSCs group-3 animal hearts as compared with EmpMSC group-2 animal hearts (magnification = 20x).