Keywords: :
In the last years, two independent concepts have improved our understanding of cancer recurrence and spread: (1) the cancer stem cell (CSC) hypothesis and (2) the occurrence of epithelial-to-mesenchymal transition (EMT). Recent evidences, such as the one presented in a recent issue by Borgna et al.,Citation1 point to a line of convergence of the two concepts.
EMT is a physiological cell reprogramming event utilized in tissue remodeling during embryonic development and activated in normal adult tissues during regeneration.Citation2 The presence of EMT-like cells in tumors has been linked to increased invasive and metastatic properties. The CSC hypothesis postulates the existence of hierarchically high-positioned, chemoresistant cells, which are responsible for disease relapse after treatment with debulking agents. These cells are endowed with the ability to reconstitute the histological heterogeneity of the originating tumor upon transplantation in immunodeficient hosts.Citation3 These properties of CSCs have strict resemblance to tissue remodeling and repair, which are typical features of mesenchymal tissues. Indeed, it has been shown that cultured breast cells that have undergone EMT in vitro also possess cancer stem cell signatures and properties.Citation4
Growth of breast cancer cells as non-adherent spheroids in relatively non-differentiating conditions is regarded as a useful tool to enrich cells endowed with CSC-like features, such as chemoresistance and tumor-repopulating ability.Citation5 In the November 2012 issue of Cell Cycle, Silvia Borgna and colleagues provide evidence, at a molecular level and by using a large panel of cell lines corresponding to different breast cancer subtypes, that mammosphere-inducing growth conditions enrich for EMT-like cell subpopulations as well.Citation1 This is especially true for Claudin-low breast cancer cell lines, which are highly enriched for CSC-like, CD44high/CD24low cells. Their work once more suggests that acquisition of EMT and CSC features are highly interconnected processes, possibly relevant for the organization of mammospheres. Indeed, recent evidence has been provided that the interaction of cell subpopulations with distinct mesenchymal and epithelial traits is instrumental for the maintenance of CSC-like cellsCitation6 and relies upon cytokine-mediated signalingCitation7 (). In light of this, it may be worth noticing that mammospheres are heterogeneous in composition and represent an ideal place for paracrine signaling to occur between different cell subpopulations. One may thus predict that compounds interfering with this crosstalk can block mammosphere formation (). Indeed Butein, a naturally occurring STAT3 and NFκB inhibitor, impairs mammosphere formation from multiple breast cancer cell lines,Citation5 possibly by blocking IL-6 signaling.Citation6
Figure 1. Schematic hypothetical working model. Mammospheres contain both CSC and EMT-like cells. When grown as mammospheres in non-adherent and no-serum conditions, breast cancer cell cultures are progressively enriched in EMT-like cells. Inset: A functional crosstalk is established between mesenchymal-like cells and epithelial-like cancer stem cells. This leads to enrichment for both CSC-like and EMT-like cell subpopulations within mammospheres. Please note that the number of CSC-like breast cells is almost constant in serially passaged mammospheres, while the proportion of EMT-like cells increases. Natural compounds, like butein, may interfere with paracrine signaling, sustaining the emergence of the mentioned cell subpopulations.
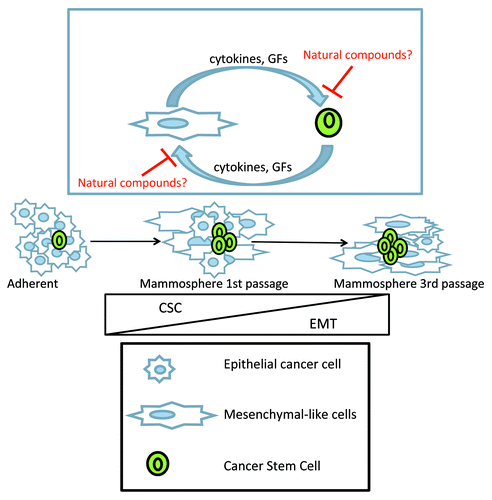
Interestingly, Borgna et al. found at least one of the known EMT-promoting transcription factors to be dynamically modulated in most of their cultures in time (mainly SNAI2 and TWIST1) when shifting from adherent to mammosphere culture conditions.Citation1 This underscores the relevance of such a process and its activation by distinct, converging and interconnected pathways. Indeed, forced expression of individual EMT-inducing transcription factors in stabilized cell lines has led Weinberg and collaborators to postulate the existence of an EMT interactome of transcription factors which are capable of reciprocally influencing each other. It will be interesting to evaluate the levels of EMT-promoting factors upon chemotherapy treatment of spheroids in vitro and to establish whether the enrichment for EMT-like cells is relevant to chemoresistance of mammospheres.
Culturing cancer cells as 3D spheroids may represent, therefore, a simplified albeit very useful tool for reproducing in vitro transient dynamic states of the tumor growth. It may also stimulate a shift in the way we envision hunting for novel therapeutic tools. Finally, this methodology is of general value, as it can be applied to cancers from other histotypes. For example, cells derived from biopsies or from malignant pleural effusions of patients with NSCLC give rise efficiently to propagating tumor spheroids in culture,Citation8 which are, again, enriched in CSC markers.Citation8,Citation9 In conclusion, in vitro cultures of tumor spheroids from stabilized cell lines and from fresh tumor specimens may therefore be considered a useful in vitro model to screen for new agents capable of co-targeting both CSCs or EMT malignant features of cancer cells.
References
- Borgna S, et al. Cell Cycle 2012; 11:4242 - 51; http://dx.doi.org/10.4161/cc.22543
- Kalluri R, et al. J Clin Invest 2009; 119:1420 - 8; http://dx.doi.org/10.1172/JCI39104; PMID: 19487818
- Alison MR, et al. Cancer Treat Rev 2012; 38:589 - 98; http://dx.doi.org/10.1016/j.ctrv.2012.03.003; PMID: 22469558
- Mani SA, et al. Cell 2008; 133:704 - 15; http://dx.doi.org/10.1016/j.cell.2008.03.027; PMID: 18485877
- Cioce M, et al. Cell Cycle 2010; 9:2878 - 87; http://dx.doi.org/10.4161/cc.9.14.12371; PMID: 20581442
- Liu S, et al. Cancer Res 2011; 71:614 - 24; http://dx.doi.org/10.1158/0008-5472.CAN-10-0538; PMID: 21224357
- Scheel C, et al. Semin Cancer Biol 2012; 22:396 - 403; http://dx.doi.org/10.1016/j.semcancer.2012.04.001; PMID: 22554795
- Mancini R, et al. PLoS One 2011; 6:e21320; http://dx.doi.org/10.1371/journal.pone.0021320; PMID: 21789168
- Bartucci M, et al. Cell Death Differ 2012; 19:768 - 78; http://dx.doi.org/10.1038/cdd.2011.170; PMID: 22117197