Abstract
Recent data demonstrated that the aberrant activity of endogenous repetitive elements of the DNA in humans can drive the expression of proto-oncogenes. This article summarizes these results and gives an outlook on the impact of these findings on the pathogenesis and therapy of human cancer.
Acknowledgements
We apologize to colleagues whose articles could not be cited because of space limitations. We thank Martin Janz for critical reading of the manuscript. Work of the authors described in this article was supported in part by grants from the Deutsche Forschungsgemeinschaft, the Berliner Krebsgesellschaft, the Wilhelm Sander-Stiftung, Leukaemia and Lymphoma Research, Cancer Research UK and Yorkshire Cancer Research.
Figures and Tables
Figure 1 Expression of CSF-1 in HRS cells depends on transcription factor NFκB. Cells of the HRS cell line L428 were transfected with 60 µg of plasmid encoding the NFκB super-repressor IκBαΔN or the respective mock control along with 10 µg pEGFP. Forty-eight hours after transfection, expression of CSF-1 was analyzed in GFP-positive cells by intracellular flow cytometry (upper part, antibody to CSF-1, MAB216; R&D Systems). IC, isotype control (MAB003; R&D Systems). Expression of IκBαΔN in L428 cells was verified by western blotting (lower part, antibody to IκBα, sc-371; Santa Cruz). Note that L428 cells lack endogenous IκBα expression due to a deleterious mutation of the NFKBIA gene.Citation55 Reh and Namalwa non-Hodgkin cell lines and expression of β-actin (antibody to β-actin, A5316, Sigma-Aldrich) were analyzed as controls.
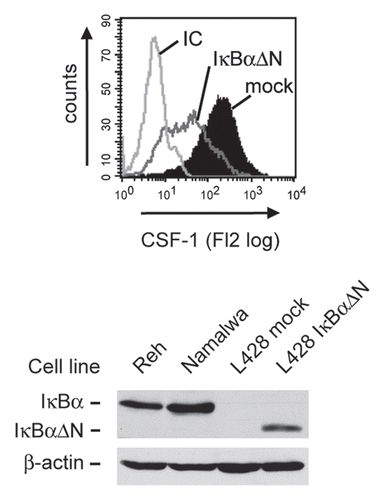
Figure 2 Model of aberrant endogenous LTR activation in Hodgkin lymphoma cells. Upper panel, In normal mature B cells, DNA upstream of the CSF1R locus including the CSF1R LTR is organized in compact heterochromatin that carries inactive histone marks such as histone H3 K9 trimethylation and methylated DNA. This recruits methyl binding proteins (MeCP), polycomb complexes (PcG) and heterochromatin protein (HP1).Citation56 The heterochromatic state of the LTR is maintained by the CBFA2T3 silencing complex. The normal CSF1R promoter is repressed by the transcription factor PAX5 which blocks the PU.1-mediated recruitment of RNA polymerase II.Citation34 Lower panel, in HRS cells, PU.1 and CBFA2T3 expression is lost, silent chromatin is not maintained and DNA is demethylated. In addition, inducible transcription factors such as NFκB and AP-1 are chronically activated and bind to the LTR. Chromatin is remodeled, leading to the recruitment of RNA polymerase II to the LTR and active transcription bypassing the normal promoter.
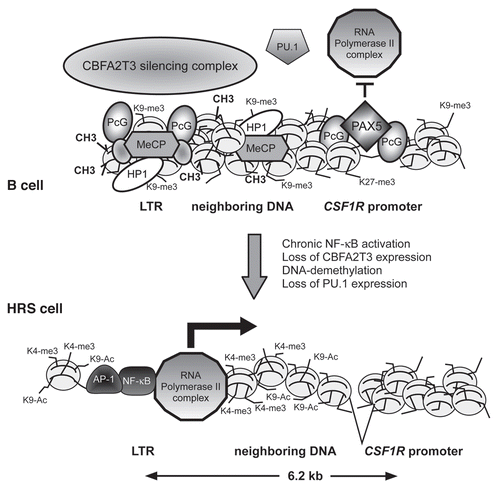
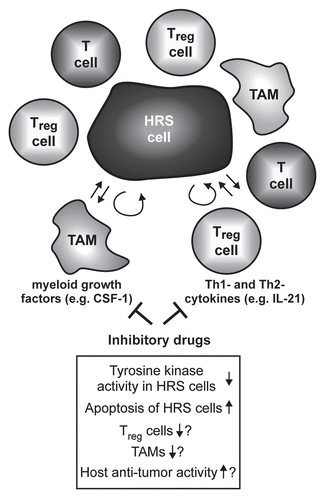
Figure 3 Simplified model of the influence of non-B cytokines on the microenvironment of HRS cells and possible consequences of their inhibition. The malignant HRS cells are surrounded by immune bystander cells like CD4+ T cells, regulatory T cells (Treg) or macrophages (among these tumor-associated macrophages, TAM). The interaction of these cells is supposed to support growth and survival of the tumor cells. Cytokines like the Th2 cytokine IL-21 or the myeloid growth factor CSF-1 are produced by the HRS cellsCitation12,Citation23 and act not only in an autocrine fashion but also paracrine on T cells and TAM. Inhibitory drugs targeting these cytokines and their associated signaling pathways might therefore not only induce growth arrest and apoptosis of the HRS cells, but also disrupt the tumor microenvironment, thereby targeting HRS cells indirectly.