Abstract
As central components of the intricate networks of eukaryotic gene regulation, transcription factors are frequent targets of ubiquitin-dependent proteolysis. A well-known example is the budding yeast MATa2 (a2) transcriptional repressor, which functions as a master regulator of cell-type determination. Degradation of a2 by the ubiquitin-proteasome system is necessary for a phenotypic switch from one cell type to another. A surprisingly complex set of ubiquitin-protein conjugation mechanisms are involved. One pathway utilizes an integral-membrane ubiquitin ligase (E3) that also functions in endoplasmic reticulum-associated degradation (ERAD). Recently, we showed that a second a2 ubiquitylation pathway uses a heterodimeric E3 that, while able to bind the ubiquitin-like protein SUMO, directly recognizes non-sumoylated a2. Other transcription factors are now also known to be ubiquitylated by multiple mechanisms; as many as a dozen E3s have been implicated in degradation of the human p53 tumor suppressor, for example. We discuss general issues of redundancy and mechanistic variation in protein modification by ubiquitin.
Acknowledgements
This work was supported by NIH grants GM046904 and GM053756 to M.H. E.M.R. was supported in part by an NRSA postdoctoral fellowship from the NIH.
Figures and Tables
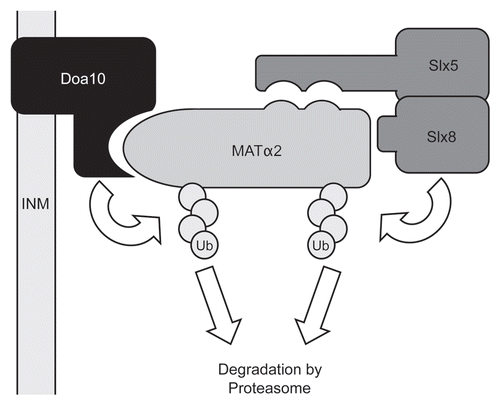
Figure 1 Ubiquitin-dependent degradation of the yeast MATα2 repressor. The ubiquitin ligases from the two major ubiquitylation pathways are represented. For clarity, ubiquitin conjugating enzymes and other accessory factors have been omitted. See text for details. Ub, ubiquitin; INM, inner nuclear membrane.