Abstract
The Ataxia-Telangiectasia mutated (ATM) kinase is regarded as the major regulator of the cellular response to DNA double strand breaks (DSBs). In response to DSBs, ATM dimers dissociate into active monomers in a process promoted by the Mre11-Rad50-Nbs1 (MRN) complex. ATM can also be activated by oxidative stress directly in the form of exposure to H2O2. The active ATM in this case is a disulfide-crosslinked dimer containing 2 or more disulfide bonds. Mutation of a critical cysteine residue in the FATC domain involved in disulfide bond formation specifically blocks ATM activation by oxidative stress. Here we show that ATM activation by DSBs is inhibited in the presence of H2O2 because oxidationblocks the ability of MRN to bind to DNA. However, ATM activation via direct oxidation by H2O2 complements the loss of MRN/DSB-dependent activation and contributes significantly to the overall level of ATM activity in the presence of both DSBs and oxidative stress.
Acknowledgements
We are grateful to the members of the Paull lab for useful suggestions. Funding was provided by the National Institutes of Health, NIH CA132813 and CA92584.
Figures and Tables
Figure 1 The FATC domain of ATM is critical for its activation. (A) A diagram of ATM domains including the FAT, PI3K kinase and FATC domains, C2991 and deletion point in the R3047X mutant allele. (B) Kinase assays were performed as described in reference Citation29, except with ATM wild-type or N2963X mutant proteins. Phosphorylation of a GST-p53 substrate was assessed by western blotting with an antibody specific for phospho-p53 serine 15 (Calbiochem, PC461). (C) An alignment of the ATM FATC domain from various species. Cysteine 2991 is marked with an asterisk. (D) The phylogram of ATM sequence alignment generated by ClustalW2. The PHYLIP method was used by ClustalW2 to generate the branching diagram. ClustalW2 phylogenetic calculations are based on the neighbor-joining method.Citation60 Branch lengths are proportional to the amount of inferred evolutionary change.
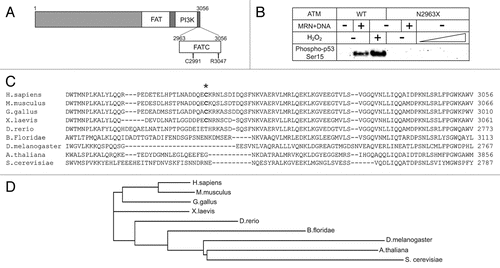
Figure 2 ATM-dependent DNA damage responses in the presence of oxidative stress. (A) Oxidative activation of ATM complements the loss of MRN/DSBs-dependent activation. Kinase assays with ATM wild-type or C2991L were performed as described in reference Citation29, in the presence of various amounts of H2O2 (0.27, 0.81 and 2.4 mM). (B) H2O2 does not affect MRN binding to ATM. Biotinylated MRN was incubated with ATM in the presence or absence of H2O2 (0.5 mM). MRN was isolated with streptavidin beads and analyzed by western blotting for Mre11 and ATM as indicated. (C) H2O2 inhibits MRN binding to linear DNA in a gel shift assay. Purified MRN was incubated with various amounts of H2O2 (34.4, 68.8, 137.5, 275 µM) before binding to a Cy5-labeled 41 bp dsDNA substrate (37°C, 10 minutes). The binding reaction was resolved on an agarose gel (0.7%, 0.5x TBE) and visualized for Cy5 fluorescence.
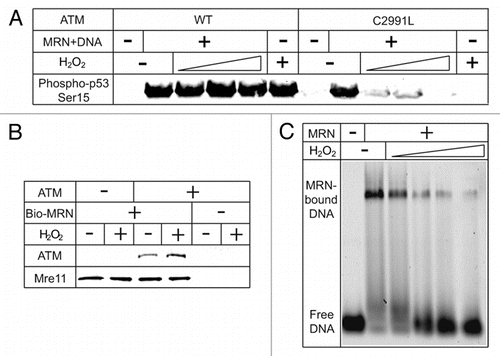
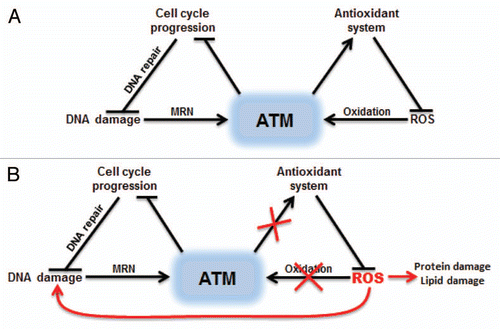
Figure 3 Dual activation of ATM by DNA damage or oxidative stress. (A) ATM can be activated by DSBs with the help of MRN complex or by direct oxidation. Oxidation of ATM is proposed to increase the activity of cellular antioxidant systems. (B) Inhibition of ATM oxidative activation results in ROS accumulation, which can damage many cellular components including DNA.