Abstract
The last universal common ancestor (LUCA) might have been either prokaryotic- or eukaryotic-like. Nevertheless, the universally distributed components suggest rather LUCA consistent with the pre-cell theory of Kandler. The hypotheses for the origin of eukaryotes are briefly summarized. The models under which prokaryotes or their chimeras were direct ancestors of eukaryotes are criticized. It is proposed that the pre-karyote (a host entity for α-proteobacteria) was a remnant of pre-cellular world, and was unlucky to have evolved fusion prohibiting cell surface, and thus could have evolved sex. The DNA damage checkpoint pathway could have represented the only pre-karyotic checkpoint control allowing division only when DNA was completely replicated without mistakes. The fusion of two partially diploid (in S-phase blocked) pre-karyotes might have represented another repair strategy. After completing replication of both haploid sets, DNA damage checkpoint would allow two subsequent rounds of fission. Alternatively, pre-karyote might have possessed two membranes inherited from LUCA. Under this hypothesis symbiotic α-proteobacterial ancestors of mitochondria might have ancestrally been selfish parasites of pre-karyote intermembrane space whose infection might have been analogous to infection of G--bacterial periplasm by Bdellovibrio sp. It is suggested that eukaryotic plasma membrane might be derived from pre-karyote outer membrane and nuclear/ER membrane might be derived from pre-karyote inner membrane. Thus the nucleoplasm might be derived from pre-karyote cytoplasm and eukaryotic cytoplasm might be homologous to pre-karyote periplasm.
Acknowledgements
We thank the Ministry of Education of the Slovak Republic (grant VEGA 1/3249/06, to Juraj Krajčovič) and Comenius University (grant UK/98/2006, grant UK/144/2007 to Matej Vesteg) for support. We wish to thank anonymous reviewers for their useful comments that contributed to the final version of the manuscript. We thank František Baluška (Institute of Cellular and Molecular Botany, University of Bonn, Bonn, Germany) for initiating us to write this manuscript.
Figures and Tables
Figure 1 Hypothetical pre-karyote cell cycle and life cycle controlled by ancient DNA damage checkpoint. The haploid pre-karyote is assumed to start DNA replication (A). If there was no DNA damage, DNA replication would be completed (B) resulting in diploid entity (C), that would be allowed to divide to produce two haploid pre-karyotes (D). If there were DNA damage (E) resulting in broken replication forks, DNA damage checkpoint would stop replication to allow the DNA repair and the restart of DNA synthesis via recombiantion with the matrix DNA (F). After completing the DNA replication (G), diploid pre-karyote (C) would be allowed to divide and the result would be two haploid entities (D). If the DNA damage were huge and pre-karyote would not be able to repair it and restart the DNA synthesis by itself (H), it could fuse with the partner (perhaps damaged too) (I). After the repair of the damage and the restart of DNA synthesis via recombination with the DNA of the partner (J), replication of both haploid sets would result in a tetraploid entity (K). Now the cell division would be allowed (L) resulting in two diploid entities. However, these would have no need to start replication again, instead both would be allowed to divide again (M and C) and the result would be four haploid entities (N and D).
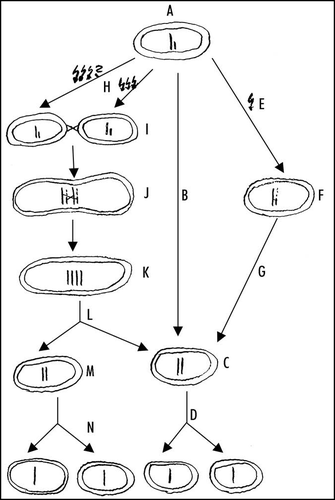
Figure 2 The hypothetical scenarios for the origin of two-membrane-bounded LUCA. (A) The fusion occurring at the orifice of a single gastruloid membrane vesicle. (B) The fusion of two cup-shaped membrane vesicles. (C) The fusion of more membrane vesicles. (D) Vesicular budding analogous to endocytic invagination. Abbreviations: V, membrane vesicle; F, the place of fusion; R, ribosomes.
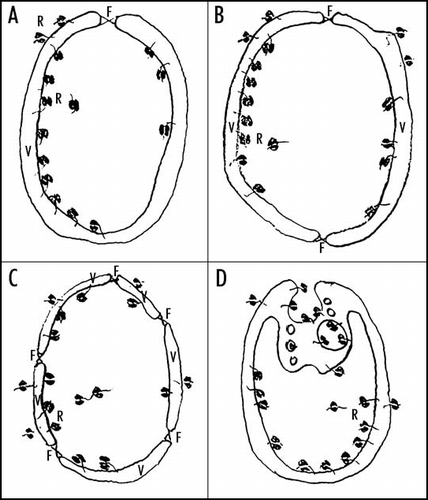
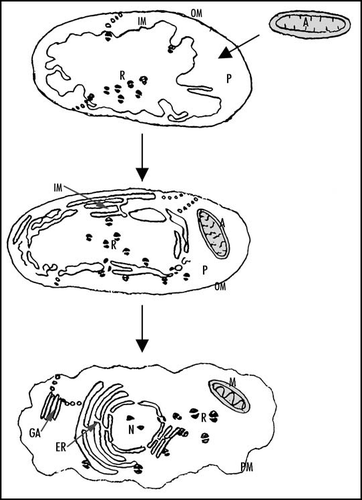
Figure 3 Hypothetical scenario for the origin of eukaryotic endomembranes. The pre-karyote is assumed to have been bounded by two membranes, inner (IM) and outer (OM). α-proteobacterial ancestors (A) of mitochondria (M) are proposed to have been parasites of pre-karyote periplasm (P). Endoplasmic reticulum (ER), nuclear membrane (N = nucleus) and Golgi apparatus (GA) are assumed to be derived from pre-karyote inner mebrane, while eukaryotic plasma membrane (PM) is assumed to be derived from pre-karyote outer membrane. It is suggested that ribosomes (R) were removed to pre-karyote periplasm (nowadays eukaryotic cytoplasm).