Abstract
For more than a century the idea that natural selection favors high levels of floral integration has prevailed as a paradigm among floral biologists. However, there is very little empirical evidence of the adaptive value of floral integration. In this addendum we highlight an important result derived from an empirical study complemented with a literature review and mathematical simulations. Results indicated that intrafloral integration but not floral integration was selected among four species of Rosaceae. The literature review coupled with null models revealed that flowering plants have on average lower than expected levels of floral integration. Mathematical simulations further demonstrated that observed levels of floral integration might result from selection favoring increased intrafloral integration. Altogether, these findings suggest that in most flowering plants, floral integration has a low adaptive value and could be a by-product of selection favoring intrafloral integration.
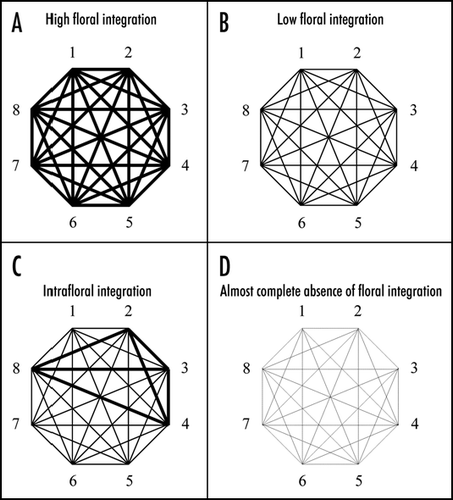
To some extent all living organisms express some degree of phenotypic integration as the result of the coordinated functions that ensures life. Thus integration itself constitutes an adaptation intimately related with the evolution of life on earth. Although empirical evidence has reported the presence of variation in the magnitude of integration among families, species, populations and individuals, few studies have examined the adaptive value of this variation (, B and D).Citation1 Because integration is directly related with functionality, the magnitude of integration can have important consequences for the interaction between species and the environment.Citation1,Citation2
In most species of Angiosperms, the flowers represent complex modular structures that evolved to ensure both reception and exportation of pollen via biotic pollination.Citation3 Because this complex function requires the morphological and behavioral matching between plants and animal vectors, the idea that high levels of floral integration are favored by natural selection was put forward since the conception of evolutionary biology ().Citation4 This hypothesis was further developed by the establishment of the pollination syndrome concept: functional and morphological characteristics of the flower in relation with the type of animal pollinator.Citation5 Moreover, Berg's concept of correlation Pleiades synthesized Darwin's ideas by postulating that, in order to ensure a correct matching between floral and pollinators morphology and behavior, species with highly specialized pollination systems should express higher levels of floral integration than those with generalists pollinators.Citation6 Thus, the Darwinian view of flowers as highly specialized mechanic devices (and therefore highly integrated structures) has become the paradigm among floral biologists.Citation7,Citation8 However, after more than a century of Darwin's ideas, there is a lack of empirical evidence of the adaptive value of the extent of floral integration. Thus, we are still far from rejecting/accepting this hypothesis given the absence of studies on the adaptive value of floral integration. Although the comparative method has provided some insights about the importance of phenotypic integration with respect to historical contingencies,Citation8,Citation9 we still do not know if natural selection promotes flower integration. On the other hand, Lande and ArnoldCitation10 have encouraged the study of natural selection with a multivariate approach, but few empirical studies have shown consistent evidence that selection favors the coordinated expression of floral attributes (e.g., significant correlational selection gradients).Citation11 Moreover, we still do not know the magnitude of the natural levels of floral integration that characterize flowering plants, and so we are unable to make contrasts with the few existing empirical studies and to determine the magnitude of integration that we should consider high or low (, B and D). Thus, in order to determine the adaptive value of floral integration we used three complementary approaches. In four species of Rosaceae (two populations each) we applied the comparative method, hierarchical selection analyses on floral integration, and performed a review of the available literature to assess the distribution of levels of floral integration among flowering plants.
Our results following the comparative method indicated that although species and clades showed significant differences in the variance-covariance structure of the flower, they present similar levels of floral integration (22.08%).Citation12 Hierarchical selection analyses on individual floral traits, scores from principal components (intrafloral integration) and the integration of the whole flower indicated that natural selection acted on individual traits and on intrafloral integration, but not upon the integration of the whole flower ().Citation12 Additionally, our review indicated that the mean level of floral integration among flowering plants is 21% of the maximum possible integration. This value resulted lower than that expected by chance (33%), supporting the idea that plants have low levels of floral integration Altogether, the three components of our study suggest a low adaptive value of floral integration.Citation12
Why Plants have Low Levels of Floral Integration?
There are at least two possible answers to this question. First, evidence has accumulated during the last decades indicating that the selection pressures exerted by pollinators may fluctuate through time, thus precluding the evolution of high levels of floral integration. In addition, several other selective agents can act upon the floral phenotype such as herbivores, florivores, nectar robbers and parasites among others.Citation13 It is common that different selective pressures act in opposite directions, potentially constraining the evolution of high levels of matching between the floral phenotype with the most important pollinator (i.e., a high level of floral integration) (). Moreover, selection acting on fruit (dispersal) traits could also constrain the evolution of high levels of floral integration because of developmental linkage, limiting the evolution of high levels of floral integration.Citation12 However, some of the most overwhelming examples of coadaptation between flowering plants and pollinators point out that the fusion of floral parts was a repeated evolutionary path followed by several lineages of plants increasing the extent of floral integration. In these cases, merging floral structures increased floral integration and is likely to represent an adaptation. Hence, besides the possible existence of constraints these cases suggest that such constraints have been overcome by different groups of species in several families.
Second, generalized low levels of floral integration could have been favored during evolutionary history, either because natural selection maintains low levels of integration on the whole variance-covariance structure of the flower, or has acted mainly to increase the integration of subsets of traits (intrafloral integration) ().Citation2 Given that we found no evidence of selection acting on the integration of the whole flower but instead promoted intrafloral integration, the low level of integration found in our review suggests that (whole) floral integration has little adaptive value. In a companion numerical simulation we additionally demonstrated that increasing intrafloral integration would indirectly augment whole flower integration, and that very low levels of intrafloral integration (30%) are sufficient to produce the observed levels of floral integration. Thus, low levels of floral integration may constitute a by-product of selection acting on intrafloral integration. This explanation suggests that relatively few floral traits mediate the interaction with pollinators, while the relationships among the rest of the floral traits may be the result of developmental/historical contingencies. If high levels of integration represent an evolutionary constraint, low levels may facilitate diversification (evolvability).
The study of phenotypic integration has become a central issue in evolutionary biology.Citation1,Citation14 Although both micro and macro evolutionary studies would provide important empirical evidence to improve our understanding of the evolution of floral integration, to date, there are probably more questions than answers to understand the adaptive value of floral integration. In this sense, our study was intended to stimulate the discussion and further research on a complex trait embedded within one of the major evolutionary novelties developed by plants, the flower.
Figures and Tables
Figure 1 Schematic representation of the different forms of association among characters in a structural or functional module. Each polygon represents a group of traits that characterize the module (i.e., flower). Line thickness indicates the degree of correlation between pairs of traits within a hypothetical module. Numbers represent characters that are components of an hypothetical module. (A) Highly integrated modules are those that present high levels of correlation between traits. (B) Modules with low levels of integration are those with weak correlations between characters. (C) Intrafloral integration indicate that a subset of characters (8, 2, 3 and 4) within the module present higher levels of correlation among them than with the rest of the characters. (D) A non integrated module can be represented by non existent or very weak correlations between characters.
Acknowledgements
This research was financially supported by a postdoctoral fellowship from the European Economic Community at the Estación Biológica de Doñana, Spain to César A. Domínguez. Mariano Ordano was supported by a posdoctoral fellowship from Universidad Nacional Autónoma de México at the Institute of Ecology. Ruben Pérez helped us during the course of this study and Etzel Garrido made useful comments on the final version of the manuscript.
Addendum to:
References
- Pigliucci M, Preston K. Phenotypic integration. Studying the ecology and evolution of complex phenotypes 2004; New York Oxford University Press
- Herrera CM. Deconstructing a floral phenotype: do pollinators select for corolla integration in Lavandula latifolia?. J Evol Biol 2001; 14:574 - 584
- Bell GB. On the function of flowers. Proc Royal Soc London B 1985; 224:223 - 265
- Darwin CR. On the various contrivances by which British and foreign orchids are fertilised by insects, and on the good effects of intercrossing 1862; London, UK Murray
- Faegri K, van der Pijl L. The principles of pollination ecology 1979; Oxford Pergamon Press
- Berg RL. The ecological significance of correlation Pleiades. Evol 1960; 14:171 - 180
- Stebbins GL. Adaptive radiation of reproductive characteristics in Angiosperms, I: pollination mechanisms. Ann Rev Ecol Evol Syst 1970; 1:307 - 326
- Armbruster SW, Pélabon C, Hansen TF, Mulder CPH. Pigliucci M, Preston K. Floral integration, modularity and accuracy: distinguishing complex adaptations from genetics constraints. Phenotypic integration. Studying the ecology and evolution of complex phenotypes 2004; New York Oxford University Press 23 - 49
- Pérez F, Arroyo MTK, Medel R. Phylogenetic analysis of floral integration in Schizanthus (Solanaceae): does pollination truly integrate corrolla traits?. J Evol Biol 2007; 20:1730 - 1738
- Lande R, Arnold SJ. The measurement of selection on correlated characters. Evol 1983; 37:1210 - 1226
- Kingsolver JG, Hoekstra HE, Koekstra JM, Berrigan D, Vignieri SN, Hill CE, Hoang A, Gilbert P, Beerli P. The strength of phenotypic selection in natural populations. Am Nat 2001; 157:245 - 261
- Ordano M, Fornoni J, Boege K, Domínguez CA. The adaptive value of phenotypic floral integration. New Phytol 2008; http://dx.doi.org/10.111/j.1469-8137.2008.02523.x
- Strauss SY, Whittall JB. Harder LD, Barrett SCH. Non-pollinator agents of selection on floral traits. Ecology and Evolution of Flowers 2006; New York Oxford University Press 120 - 138
- Murren C. Phenotypic integration in plants. Plant Species Biol 2002; 17:89 - 99