
Abstract
Symmetric cell division in Gram-negative bacteria requires the concerted action of three Min proteins that together ensure exclusive formation of the cell division septum at the mid-point of the cell. We have recently described the structure and dynamic properties of MinE, the protein responsible for directing the cell division inhibitor complex formed by the MinC and MinD proteins away from the middle of the cell. An unexpected feature of this structure was the location of MinD-binding residues at buried, non-accessible sites in the dimeric interface. Here we elaborate on the potential role of conformational changes that might be involved to allow access to these residues, along with the interesting questions raised by these features of the MinE structure.
Figures and Tables
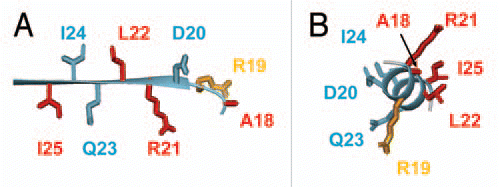
Figure 1 (A) Two views of the solution NMR structure of MinE from Neisseria gonorrhoeae (PDB accession number 2KXO), with a model of the more open conformation suggested by NMR dynamics data on the right hand side. Each subunit in the dimer is shown in a different colour, and β-strands β1, β2 and β3 labeled for each subunit. Sidechain atoms for residues that directly participate in MinD interactions are also shown, with the inaccessible Leu-22 highlighted in red and the partially inaccessible residues (Arg-21 Ile-25) shown in orange. (B) The solution NMR structure of residues 31–88 from E. coli MinE (1EV0), showing the different dimer interface formed by residues corresponding to β3 in the full-length structure.
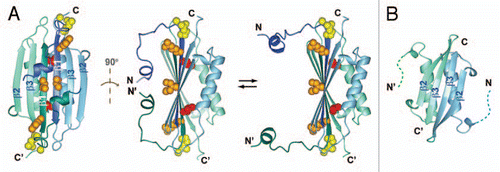
Figure 2 Magnified view of the region of MinE containing residues that bind to MinD, either in (A) the β1 structure as it appears in the dimer or (B) in a hypothetical α-helical structure. Sidechains that are critical for MinD binding are shown in red, those that play a smaller role in the interaction in orange and those that can be mutated and still give rise to wild-type function are in blue.
Addendum to: