
Abstract
Prestin, which is a member of the solute carrier 26 anion transporter family (SLC26A5), is a voltagedependent membrane-based motor protein that confers electromotility on mammalian cochlear outer hair cells (OHCs).1 OHCs are a mammalian innovation, their presence2 and their endowment with functional prestin is essential for normal hearing of mammals.3 In order to clarify the molecular mechanism underlying the voltage-dependent motility of prestin, precise description of the relation between voltage-induced prestin-associated charge movement and the resulting cell displacement is essential. By simultaneously measuring voltage-dependent charge movement, which is manifested in the nonlinear capacitance (NLC) of the cell membrane, and voltage-induced OHC displacement, we provided compelling experimental evidence that prestin-associated charge movement and the resulting electromotility are fully coupled, and that prestin has at least two voltage-dependent conformational transition steps. These findings provide a basis for understanding the molecular mechanism of prestin. Here we discuss the relevance of our finding in the elucidation of the voltage-dependent motor mechanism of prestin, and speculate about possible voltage sensing mechanisms of the molecule.
Acknowledgments
This work was supported by the Hugh Knowles Center and National Institutes of Health Grant DC00089-40 (to P.D.).
Figures and Tables
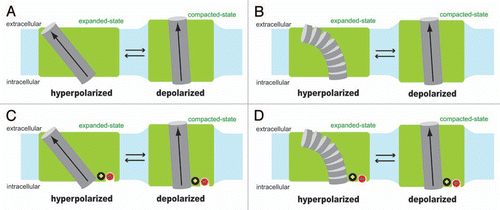
Figure 1 Hypothetical voltage sensing mechanisms of prestin. Two different structural states of prestin (expanded vs. compacted) are shown in green with a hypothetical voltage sensing α-helix (gray cylinder), whose N-terminus faces the extracellular side. The α-helix dipole, the direction of which is from the C-terminus (negatively charged) to the N-terminus (positively charged) by definition, is shown with arrow. Shown in light blue is the cell membrane. The length of the hypothetical voltage sensing α-helix is assumed to be similar to the thickness of the cell membrane in the models. If the typical cell membrane thickness of ∼3 nm were used, and if the partial charge present at the N-terminus and the C-terminus were assumed to be +0.5 and −0.5, respectively, the magnitude of the α-helix dipole moment would be calculated as 2.4 × 10−28 Cm (computation: 0.5 × 1.6 × 10−19 × 3 × 10−9). In model (A and C), the structure of the hypothetical voltage sensing α-helix is assumed to be maintained during physiological membrane potential changes, whereas, in (B and D), the propensity of the α-helix formation is assumed to be moderate or low, and thus is assumed to be significantly reduced to induce destruction of the helix structure under hyperpolarized membrane potential. In (C and D), a hypothetical intrinsic positive charge (black circle) and a hypothetical extrinsic anion binding site with bound chloride (red circle) are included for explaining the observed Vpk shifts induced by chloride substitution with various anions (see text for detail).
Addendum to: