
Abstract
During regeneration, lost structures are rebuilt and perfectly integrated within the remaining non-injured tissues. This fascinating process captured the attention of one of the founders of modern genetics, T. H. Morgan. He was particularly interested in understanding regeneration in freshwater planarians, which can regenerate a whole animal from a small piece of their bodies. He performed numerous experiments to understand how polarity is re-established such that an anterior-facing wound regenerates a head whereas a posterior-facing wound regenerates a tail. However, it has not been until more than 100 years later that the molecules required to determine axial polarity have been identified. Several studies have now shown that the Wnt/β-catenin and Hedgehog pathways are required for anteroposterior axis specification, whereas the establishment of the planarian dorsoventral (DV) axis relies on the Bone Morphogenetic Protein (BMP) pathway. Two recent papers have now uncovered additional conserved (anti-dorsalizing morphogenetic protein) and novel (noggin-like genes) elements that regulate planarian DV axis regeneration. Here, we summarize those results and present new data and hypotheses to explain the role that noggin-like genes might play.
Acknowledgments
We thank Iain Pattern for advice on English style in a version of the manuscript. M.D.M. was funded by an FPU fellowship from the Ministerio de Educación y Ciencia (Spain). F.C. is a Ramón y Cajal researcher (Ministerio de Ciencia e Innovación, Spain). This work was funded by grants BFU2008-01544 (MICINN) to E.S., grant BFU2008-00710 (MICINN) to F.C., and grant 2009SGR1018 (AGAUR, Generalitat de Catalunya) to E.S. and F.C.
Figures and Tables
Figure 1 Graphical representation of a proposed gradient of BMP signaling in the dorsal region of the planarian and its hypothetical reduction after different RNAi knockdowns. After nlg8 silencing the decrease in BMP signaling around the midline would not be sufficient to affect DV patterning. However, in the lateral regions, the reduction of BMP activity would eliminate the antineurogenic effect of the pathway and allow the differentiation of an ectopic CNS (arrows). Only after bmp silencing, alone or in combination, would the reduction in the level of BMP activity around the midline result in ventralized planarians.
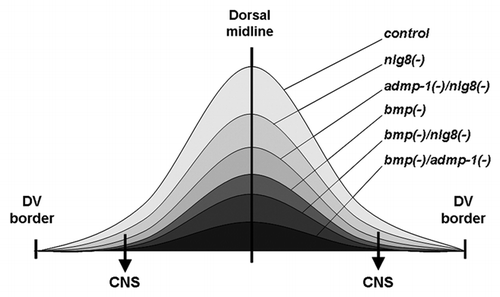
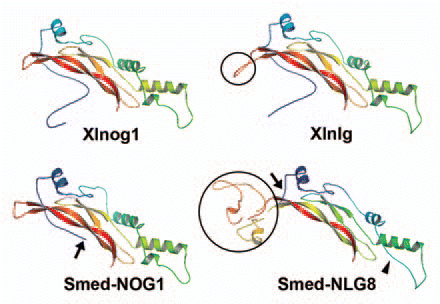
Figure 2 Comparison of the predicted protein structures of noggin and noggin-like proteins. Homology-based protein structures of Xenopus noggin1 (Xlnog1), planarian NOGGIN1 (Smed-NOG1), Xenopus noggin-like (Xlnlg) and planarian NOGGIN-LIKE 8 (Smed-NLG8) constructed by the Swissmodel programCitation17 and further modified using PyMOL software (DeLano Scientific LLC, pymol.sourceforge.net/) are shown. The structure of planarian NOGGIN shows a high degree of similarity to the Xenopus noggin1 homolog, although its BMP-binding domain is shorter (arrow). With the exception of the insertion (black circles), the predicted structures of the noggin-like homologs resemble those of noggins. Although the length of the amino-acid insertion varies between species,Citation4 its presence could probably modify the binding capacities of noggin-like homologs. Note that a helix is absent in Smed-NLG8 (arrowhead). Accession numbers: AAT91717, ABV04323, NP_001089147, ACO06233.
Addendum to: