
Abstract
Nitric oxide (NO) is an endogenous second messenger which acts as a potent vasodilator, anti-inflammatory, anti-thrombotic, and pro-angiogenic agent in the vasculature. Recent studies revealed that the effects of NO on blood vessels are mediated in part by its ability to regulate protein trafficking machinery and vesicle-based exocytosis in vascular cells. Specifically, NO-dependent S-nitrosylation of N-ethylmaleimide sensitive factor (NSF), an ATPase that enables membrane fusion, was shown to inhibit exocytosis of vesicular secretory compartments such as endothelial Weibel-Palade bodies, platelet alpha granules, and cytolytic granules from activated lymphocytes. Tissue transglutaminase (tTG or TG2) is a multifunctional protein synthesized and secreted by various cell types in the vasculature, which is involved in multiple vascular diseases, including atherosclerosis, vascular calcification, and age-dependent aortic stiffening. Our recent findings indicate that tTG is delivered to the cell surface and the extracellular matrix (ECM) via a non-classical ER/Golgi-independent secretion pathway, which depends on the recycling endosomes and the NSF function. Here we report that NO attenuates the unconventional secretion of tTG in human aortic endothelial cells. NO-dependent down-regulation of extracellular tTG levels via inhibition of its secretion might be a part of general physiological mechanism which limits externalization of adhesive, pro-inflammatory and thrombogenic proteins in the vasculature.
Acknowledgments
This work was supported by NIH grant 2RO1 HL105296 to D.E.B.
Figures and Tables
Figure 1 Effects of NO on externalization of endogenous tTG in HAECs. (A) Untreated cells and cells pretreated with NO donor GSNO (200 µm, 1 h) or NOS inhibitor L-NAME (100 µm, 18 h), were metabolically labeled with 35S-translabel for 0–8 h. at the end of labeling, cells were surface-biotinylated with Sulfo-NHS-LC-biotin and cell surface protein fractions were isolated on neutravidinagarose.Citation5,Citation7 the de novo synthesized tTG was immunoprecipitated from total cell extracts (left parts) and cell surface protein fractions (right parts). the resulting tTG immune complexes were resolved by SDS-PAGE and visualized by fluorography. Large and small arrowheads mark the positions of tTG (∼80 kDa) and its major proteolytic fragment (∼68 kDa). (B) the relative amounts of de novo synthesized cell surface tTG were quantified by scintillation counting and compared to those of total tTG. Shown in (A) is a representative of four independent experiments. Bars in (B) depict means ± Sem. *p < 0.05.
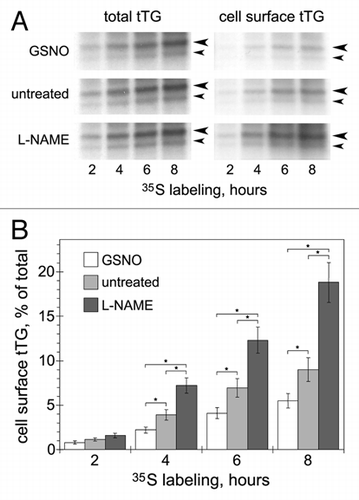
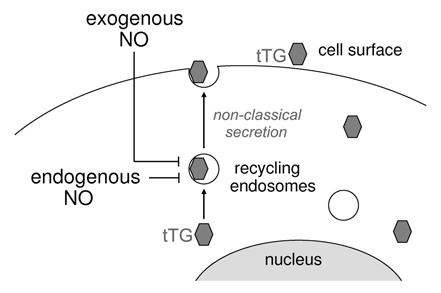
Figure 2 A scheme depicting the effects of exogenous and endogenously produced NO on nonclassical secretion of cytoplasmic tTG.
Addendum to: