
Abstract
Synaptic transmission is a major mechanism by which neurons communicate with each other. Basic steps in neurotransmitter release are similar in all synapses. However, many properties of release vary between synapses and reflect specific structural and functional requirements, endowing synapses with specialized functions. Recently, Gelman et al.1 described properties of release and short-term depression at specialized nicotinic synapses in the brainstem of goldfish, Carassius auratus (Linnaeus). These axo-axonic synapses between the Mauthner cell collaterals and their targets, cranial relay neurons (CRNs), exhibit strong short-term depression, even at stimulation frequencies as low as 0.33 Hz. In short, amplitudes of post-synaptic responses, evoked by pre-synaptic trains of action potentials, were depressed with a time course approximated by a sum of two exponential functions. Initially, fast depression reduced the amplitude of EPSP2 (response after the second stimulus), to less than 50% of EPSP1 (response after the first stimulus). This was followed by a slow component of depression that produced an additional 10-30% amplitude reduction over a time-span of tens to hundreds of seconds. Interestingly, depressed EPSPs exhibited longer latencies than that of the ‘undepressed’ EPSP1. Additionally, fast and slow calcium chelators (BAPTA and EGTA), injected pre-synaptically, were equally potent in reducing release. These data are consistent with a previously proposed general mechanism that assumes a change in release probability after the initial release. However, in an alternative interpretation the results could be coherently explained by postulating two releasable pools of vesicles, with high and low release probabilities, and a generally accepted depletion scheme. This latter interpretation will be discussed in this article.
Acknowledgments
Original research was conducted in the laboratory of Dr. Donald S. Faber (Albert Einstein College of Medicine). The support was provided by NIH training grant T32NS007439.
Figures and Tables
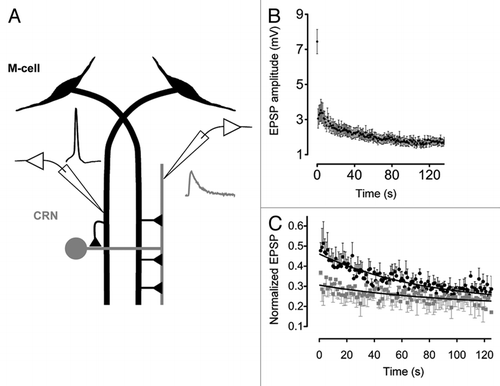
Figure 1 (A) A diagram depicting the M-cell network and recording arrangement. Traces depict M-cell spike (black) generated by injecting depolarizing current and evoked post-synaptic EPSP (gray). (B) EPSP amplitudes during a 1 Hz stimulus train (n = 19 different pairs; mean ± SEM ). (C) Ca2+ buffer's effect on the kinetics of depression. Black circles and gray squares are normalized EPSP amplitudes (EPSP1 is omitted) evoked by pre-synaptic trains of stimulation (1 Hz) before and after injection of 10 mM BAPTA. Black curves are fitted single exponential functions.
Addendum to: