Abstract
The retromer complex is conserved across all eukaryotic species and functions in physiologically important sorting processes at the endosomal membrane. A key component of retromer is the VPS29 protein that, although structurally similar to phospho-diesterases, has been convincingly shown in the recent study by Swarbrick et al. (PLoS One 6:e20420, 2011) to be a rigid scaffold that interacts with various proteins that function with retromer in endosomal protein sorting. A widely held view, based on initial observations in Saccharomyces cerevisiae, is that retromer functions as a stable heteropentamer. This is, however, contrary to experimental data presented in Swarbrick et al. (and in other studies) that indicate that retromer in higher eukaryotes is a looser association of two subcomplexes that respectively mediate cargo-selection and membrane tubulation. Here we present an analysis of evolutionary variation of the VPS29 protein and discuss why the retromer complex as first characterized in S. cerevisiae is not representative of retromer in other eukaryotic taxa.
The retromer complex is a key mediator of endosomal protein sorting and trafficking from endosomes to the Golgi complex. Recently there have been a number of studies implicating retromer in diverse physiological processes that involve recycling of receptors and ion channels and therefore there is much interest in understanding the molecular mechanisms of retromer function.Citation1,Citation2
Retromer was first identified and described in the budding yeast, Saccharomyces cerevisiae comprising five proteins that operate together at the endosome membrane. Although retromer functions as a heteropentamer in yeast, phenotypic and biochemical studies enabled the complex to be functionally dissected into two subcomplexes; the cargo-selective subcomplex is a trimer of the VPS35, VPS26 and VPS29 proteins, whilst the SNX-BAR dimer of the Vps5 and Vps17 proteins act to form membrane tubules at the endosome.Citation3 The VPS29 protein links the two subcomplexes together with a conserved hydrophobic patch on the surface of VPS29 being critical for the assembly of the heteropentamer. In mammals and other higher eukaryotes, the SNX-BAR proteins, SNX1 and SNX2 serve in place of Vps5p whilst Vps17p function is performed by SNX5 and SNX6 (see ).
Species-Specific Variations
In the recent study of Swarbrick et al.Citation4 the authors reaffirm that the phosphodiesterase-like fold of VPS29 is not enzymatically active and that VPS29 does not undergo any significant structural changes between free and complexed states that could create such activity. Additionally the authors also show that the mammalian VPS29-SNX1 interaction reported previously is, though specific, in fact very weak, and conclude would be insufficient in vivo for complex formation without further factors and interactions. This is in contrast to what might have been expected from the model-forming experiments performed in S. cerevisiae but is consistent with data from native immunoprecipitation experiments performed using lysates from mammalian cells.
Given that retromer is conserved across all eukaryotes,Citation5 it is perhaps surprising that studies of retromer in mammalian cells revealed that (unlike S. cerevisiae) the complex is not a stable heteropentamer. The cargo-selective trimer is a stable complex, as is the SNX-BAR dimer but the two subcomplexes associate only transiently. Another important difference between retromer in S. cerevisiae and mammals is that mammalian retromer associates with a number of accessory proteins including the WASH complex and a RAB GTPase activating protein (GAP), TBC1D5.Citation6 The difference between retromer in S. cerevisiae and higher eukaryotes is intriguing but also important for understanding the molecular mechanisms of retromer-mediated endosomal protein sorting.
How Representative of all Eukaryotes is Saccharomyces cerevisiae Vps29p?
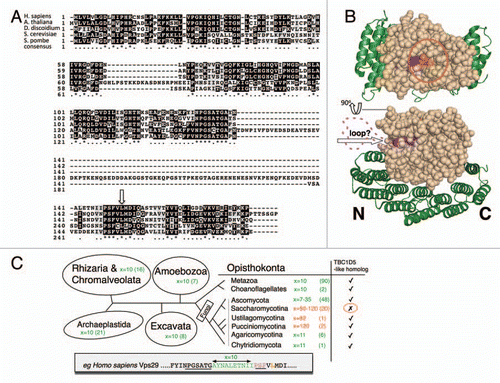
In S. cerevisiae Vps29p binds to Vps5p, in mammals VPS29 binds TBC1D5. These interactions require the conserved leucine at positions 252 and 152 respectively.Citation6,Citation7 A key difference between yeast and mammalian VPS29 is a ∼80 amino acid loop present in yeast Vps29p close to the critical leucine that forms the interaction site for Vps5p and TBC1D5 (see ). A clear implication of the presence and location of the loop is that this additional sequence may be responsible for the observed differences in the interactions of yeast and mammalian VPS29.
Expanding our analyses to additional species we have retrieved and aligned VPS29 protein sequences, and have identified significant stretches of extra residues in a subset of homologues. In Saccharomycotina (the fungal clade that includes S. cerevisiae) the Vps29 protein contains an additional protein sequence of variable length (50–120 amino acids) that is inserted at an identical site in the protein. In S. cerevisiae the insert codes for an unstructured loop of ∼80 largely hydrophilic charged amino acids. When mapped onto the known structure of VPS29 (shown in bound to a portion of VPS35), the insertions occur at a site distinct from either the conserved accessory protein interaction site or the site of VPS35 binding. The large inserted sequences were not present in other eukaryotes with the exception of a small number of Basidiomycetes in the Ustilagomycotina and Pucciniomycotina clades (see ).
How did the insertion present in Saccharomycotina VPS29 occur? We suggest that splice-site mutations in the VPS29 gene incorporated previously intronic material into the reading frame. In support of this idea we have identified appropriately located splice junctions in the VPS29 gene from the extant chytrid Batrachochytrium dendrobatidis (NCBI accession EGF82008.1), a species believed to be similar to the ancient common fungal ancestor. We therefore hypothesize that the material gained has been responsible for the greater affinity observed with Vps5p in S.cerevisiae.
A Further Correlation
In mammals, VPS29 binds to TBC1D5. TBC1D5 is not, however, uniformly conserved in evolution. The presence of a TBC1D5 homologue in a given genome may be scored by the identification of a molecule with a following features: (1) a RAB GAP (TBC) domain in the N-terminal half, (2) a second folded domain with predicted coiled-coil secondary structure, (3) ∼700–800 residues in length and (4) strongest homology to human TBC1D5 using a simple BLAST alignment. We noticed that (with very few exceptions) the species in which VPS29 contains the additional protein sequence at the site close to the critical leucine do not appear to have retained a TBC1D5 gene and vice versa. We therefore propose a causal link between these observations. The insertion of additional hydrophilic amino acids close to the key leucine residue may have resulted in a gain of function with respect to the VPS29-SNX-BAR interaction but a concomitant loss of interaction with TBC1D5. The hydrophilic nature of the inserted sequences may promote the binding of the SNX-BAR protein through charge-based interactions. The loss of the VPS29-TBC1D5 interaction may subsequently have led to the loss of the TBC1D5 gene in those fungal species where VPS29 gained the additional protein loop.
Conclusions
In light of these correlations and also the emerging reports of cargo-recognition-subcomplex-independent roles for SNXBAR heterodimers,Citation10,Citation11 we suggest that the original, yeast-derived, SNX-BAR-containing heteropentameric model of the retromer complex is not valid for the vast majority of eukaryotic taxa. The association of the cargo-selective trimer with the membrane tubulating SNX-BAR complex will likely be too weak to enable a stable heteropentameric complex to be readily identified. That does not mean however that the cargo-selective trimer and respective SNX-BAR proteins do not function together in the same pathway. To determine the prevalence of the heteropentameric retromer complex, additional experiments to analyze the composition and formation of the retromer complex in eukaryotes where VPS29 contains the additional amino acid sequences will be required.
Abbreviations
| Vps | = | vacuole protein sorting |
| SNX | = | sorting nexin |
| BAR | = | Bin-amphiphysin-Rvs domain |
| TBC | = | Tre2-bub2-Cdc16 domain |
Figures and Tables
Figure 1 Schematic diagram of the retromer complex and its role in endosome-to-Golgi retrieval. Retromer in S. cerevisiae is a stable heteropentamer comprising the cargo-selective VPS35-VPS26-VPS29 trimer and the SNX-BAR dimer of VPS5-VPS17. VPS29 links the two subcomplexes together.
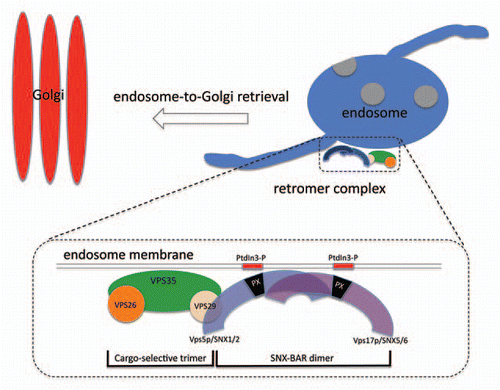
Figure 2 (A) Sequence alignment of VPS29 protein sequences from H. sapiens, A. thaliana, D. discoidium, S. pombe and S. cerevisiae. The conserved regions are shaded black. There is a large unconserved loop in the S. cerevisiae sequence that is not present in VPS29 from other species. The critical leucine involved in VPS29 interactions is highlighted with an arrow. (B) The additional loop is shown mapped onto the structure of mouse VPS29 (beige) in complex with the C-terminal domain of VPS35 (green) (based upon Hierro et al.Citation8). The loop lies close to the critical leucine residue (shown in orange) required for VPS29 to interact with Vps5 or TBC1D5. The purple shaded residues correspond to the conserved sequence P-S-F that is located just after the loop sequence. (C) VPS29 ancestral state. A survey of approximate distances (x, amino-acid residues) between the conserved PGSxTG and PSF motifs reveals that large sequence insertions are derived features in certain fungal clades (number of representatives considered in parentheses). Diagram adapted from Walker et al.Citation9 (see for further phylogenetic information). A TBC1D5 homolog was identified in all of the various eukaryotes except for Saccharomycotina.
Acknowledgments
We thank Brett Collins for helpful comments. MEH and MNJS are funded through an MRC Senior Research Fellowship awarded to MNJS.
Addendum to:
Related Research Data
References
- McGough IJ, Cullen PJ. Recent advances in retromer biology. Traffic 2011; 12:963 - 971
- Temkin P, Lauffer B, Jäger S, Cimermancic P, Krogan NJ, von Zastrow M. SNX27 mediates retromer tubule entry and endosome-to-plasma membrane trafficking of signalling receptors. Nat Cell Biol 2011; 13:717 - 723
- Seaman MN, McCaffery JM, Emr SD. A membrane coat complex essential for endosome-to-Golgi retrograde transport in yeast. J Cell Biol 1998; 142:665 - 681
- Swarbrick JD, Shaw DJ, Chhabra S, Ghai R, Valkov E, Norwood SJ, Seaman MN, Collins BM. VPS29 is not an active metallo-phosphatase but is a rigid scaffold required for retromer interaction with accessory proteins. PLoS One 2011; 6:e20420
- Koumandou VL, Klute MJ, Herman EK, Nunez-Miguel R, Dacks JB, Field MC. Evolutionary reconstruction of the retromer complex and its function in Trypanosoma brucei. J Cell Sci 2011; 1241496 - 1241509
- Harbour ME, Breusegem SY, Antrobus R, Freeman C, Reid E, Seaman MN. The cargo-selective retromer complex is a recruiting hub for protein complexes that regulate endosomal tubule dynamics. J Cell Sci 2010; 123:3703 - 3717
- Collins BM, Skinner CF, Watson PJ, Seaman MN, Owen DJ. VPS29 has a phosphoesterase fold that acts as a protein interaction scaffold for retromer assembly. Nat Struct Mol Biol 2005; 12:594 - 602
- Hierro A, Rojas AL, Rojas R, Murthy N, Effantin G, Kajava AV, Steven AC, Bonifacino JS, Hurley JH. Functional architecture of the retromer cargo-recognition complex. Nature 2007; 449:1063 - 1067
- Walker G, Dorrell RG, Schlacht A, Dacks JB. Eukaryotic systematics: a user's guide for cell biologists and parasitologists. Parasitology 2011; 15:1 - 26
- Pourcher M, Santambrogio M, Thazar N, Thierry AM, Fobis-Loisy I, Miège C, Jaillais Y, Gaude T. Analyses of sorting nexins reveal distinct retromer-subcomplex functions in development and protein sorting in Arabidopsis thaliana. Plant Cell 2010; 22:3980 - 3991
- Nisar S, Kelly E, Cullen PJ, Mundell SJ. Regulation of P2Y1 receptor traffic by sorting Nexin 1 is retromer independent. Traffic 2010; 11:508 - 519