
Abstract
We recently reported evidence implicating fatty-acid binding protein (Fabp) in the control of sleep and memory formation. We used Drosophila melanogaster to examine the relationship between sleep and memory through transgenic overexpression of mouse brain-Fabp, Fabp7, or the Drosophila Fabp homologue, (dFabp). The key findings are that, (1) a genetically induced increase in daytime consolidated sleep (naps) correlates with an increase in cognitive performance, and (2) a late “window” of memory consolidation occurs days after the traditionally understood “synaptic” consolidation. Exactly how Fabp-signaling may be involved in converting normal to enhanced long-term memory (LTM) is not known. Here we describe additional data which support relative subcellular compartmental localization of Fabp in regulating stage associations of different forms of memory in Drosophila. Anesthesia resistant memory (ARM) is a longer lasting memory that is produced by massed training, but unlike LTM produced by spaced training, it is insensitive to protein synthesis inhibitors and does not persist as long. We observed that the ratio of ARM to LTM performance index of Fabp7-transgenic flies is proportional to the relative cytoplasmic to nuclear Fabp7 expression level. These data suggest a common lipid-signaling cascade exists between phases of memory formation previously thought to be molecularly distinct.
Acknowledgments
We would like to thank H. Zhou for expert technical assistance with Westerns, and A. Pack and the Center for Sleep and Circadian Neurobiology for support on this manuscript. J.R.G. is currently supported by NIH T32 HL07713.
Figures and Tables
Figure 1 Fabp7 flies have enhanced ARM formation. Fabp7 flies elicit Anesthesia Resistant Memory (ARM) enhancement following 10X Massed (10XM) training when maintained at lower (20°C) and higher temperatures (30°C) for 4 days prior to testing, compared to controls. Grey bars, Fabp7, open bars, w(isoCJ1) background strain. Results are mean ± s.e.m.; n = 8 groups. * P < 0.05, t-test Fabp7 vs. w(isoCJ1).
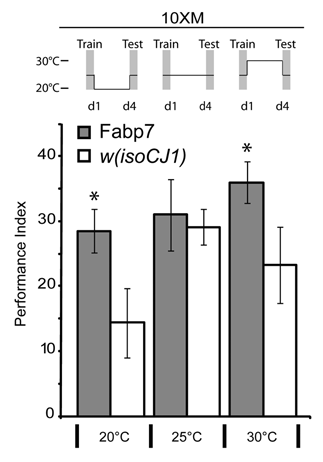
Figure 2 Subcellular localization of Fabp7 correlates with temperature. Protein gel blots of Fabp7 protein expression in Fabp7 fly heads staged at 20°, 25°, and 30°C for seven days, compared to immunoreactive background, in cytoplasmic and nuclear fractions previously publishedCitation8 (lysate sample: ∼50–100 fly heads per lane). Image contrast has been inverted for comparison.
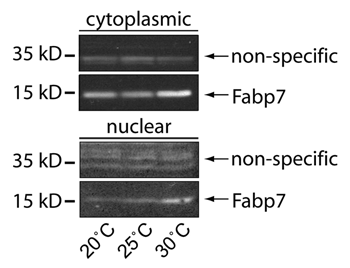
Figure 3 Relative subcellular localization of Fabp7 correlates with memory formation. (A) Ratio of Performance Index (PI) scores from ARM data (), to Long-Term Memory following (10X Spaced) training previously described.Citation8 Grey bars, Fabp7, open bars, w(isoCJ1) background strain. Results are mean ± s.e.m.; n = 8–10 groups. * P < 0.05, t-test, vs. Fabp7 at 25°C. (B) Ratio of Performance Index (PI) scores in () for Fabp7 flies (diamonds; y-axis left) shows a correlation of performance with ratio of cytoplasmic to nuclear (Cyto/Nuc) Fabp7 protein expression (black circles; y-axis right; from ).
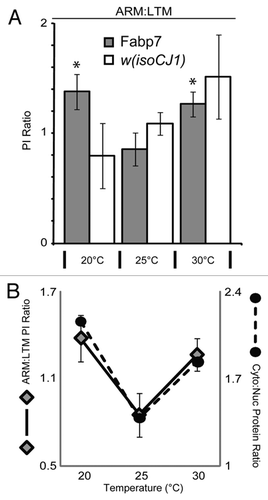
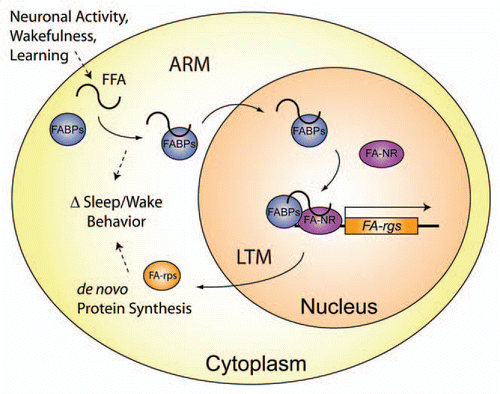
Figure 4 Proposed model for Fabp cellular processes regulating sleep and memory formation. Upon neuronal activation, wakefulness, or learning events (such as repeated stimulation during ARM training), free fatty-acids (FFA) are liberated from the cell membrane or other lipid stores, and can modulate sleep/wake. Increases in the relative amount of cytoplasmic Fabps bind more FFA and thereby regulate cellular responses to lipid signals. Following LTM training, additional FFA are liberated, to which Fabp will sequester and transport to the nucleus in order to activate fatty-acid nuclear receptors (FA-NR) and regulate transcription of downstream fatty-acid responsive genes (FA-rgs). This altered transcription will generate a change in protein synthesis of FA-rgs (FA-rps), which in turn modulates sleep/wake behavior.
Addendum to: