Abstract
The role of iron in brain physiology has focused on the neuropathological, effects due to iron-induced oxidative stress. However, our recent work has established a physiological relationship between the iron-mediated oxidative modification and normal neuronal function. Our results obtained from hippocampal neurons, suggest that iron-generated reactive species oxygen (ROS) are involved in calcium signaling initiated by stimulation of NMDA receptors. This signal is amplified by ryanodine receptors (RyR), a redox- sensitive calcium channel, allowing the phosphorylation and nuclear translocation of ERK1/2. Furthermore, using electrophysiological approaches, we showed that iron is required for basal synaptic transmission and full expression of long-term potentiation, a type of synaptic plasticity. Our data combined suggest that the oxidative effect of iron is critical to activate processes that are downstream of NMDAR activation. Finally, due to the high reactivity of DNA with iron-generated ROS, we hypothesize an additional function of iron in gene regulation.
Due to its ability to accept or transfer electrons, iron participates in a series of redox reactionsCitation1,Citation2 such as the Fenton reaction that generates hydroxyl free radicals or the Haber-Weiss reaction that combines the reduction of Fe3+ by superoxide plus oxygen to produce Fe2+.
When iron accumulates, it can promote oxidative stress which in turn triggers neuronal death.Citation3 As a result of iron-mediated reactions, the study of iron in the brain has been focused on its neuropathological role.Citation4,Citation5 However, our recent work has established a physiological relationship between the iron-mediated oxidative modification and normal neuronal function.Citation6 This work also suggests that iron-generated reactive species oxygen (ROS) could be a new class of molecules that act as second messengers in signaling cascades related to synaptic plasticity (SP), the putative cellular substrate of memory.
Consistent with a potential physiological role of iron, the activation of N-Methyl-D-aspartate (NMDA) receptors (NMDAR) induces iron uptake in cultured cortical neurons (), which in turn induces the production of the hydroxyl free radical- mediated Fenton reaction.Citation2 Similarly, in cultured hippocampal neurons, we showed that the entry of iron rapidly increased labile iron for the Fenton reaction.Citation6
Figure 1. Under basal condition (), glutamatergic synapses in the hippocampal CA1 region is coupled to proteins required to induce synaptic plasticity (SP), such as N-Methyl-D-aspartate receptors (NMDAR) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR), divalent metal transporter 1 (DMT1, Nramp1), neuronal NADPH oxidase and a large number of kinases that are activated by calcium (e.g.: ERK 1/2). At hippocampal dendrites, the endoplasmic reticulum, an intracellular calcium reservoir containing the ryanodine receptors (RyR), which are activated by calcium and reactive oxygen species (ROS). In the nucleus, regulatory sequences within the genes involved in SP (CpG islands), are frequently methylated by DNA methyltransferases (DNMTs). Methylation of cytosine at position 5, leads to the formation of 5-methyl-cytosine (5mC) which is recognized by methyl CpG binding protein 2 (MeCP2) preventing the binding of transcription factors activated by calcium, such as cAMP response element-binding (CREB). Neuronal activity () triggers critical events for expression of SP, such as insertion of AMPAR to the synapse; superoxide generation and subsequent dismutation to hydrogen peroxide; iron uptake that promotes oxidative reactions; NMDAR activation that allows the calcium influx from the synaptic cleft, which coupled with redox modifications of RyRs, amplifies the signal of calcium in a process known as calcium-induced calcium release. This process activates calcium signaling pathway such as ERK/MAPKs that phosphorylate the transcription factor CREB, regulating genes involved in SP. Iron may modulate the binding of these transcriptional regulators, forming adducts such as 8-hydroxyguanosine (8OHG) or 5 hydroxymethylcytosine (5HmC).
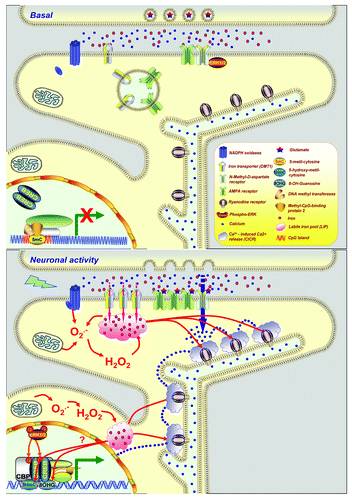
Why are there different mechanisms to incorporate iron, after neuronal activity? We propose that iron uptake is needed to provide a neuronal oxidative tone necessary for the proper functioning and operation of components sensitive to this tone. In agreement with this idea, ryanodine receptors (RyR), a channel involved in calcium- induced calcium release (CICR) which activates various signaling pathways involved in synaptic plasticity,Citation7 has a group of oxidation-sensitive cysteinesCitation8 that could be one of the putative targets of iron-mediated oxidative attacks, as proposed by earlier
work in PC12 cells.Citation9 Our latest work in hippocampal neurons suggests that iron-generated ROS are involved in calcium signaling initiated by stimulation of NMDA receptors and amplified by the RyR.Citation6 An iron chelator, deferrioxamine, prevents CICR as well as the phosphorylation and nuclear translocation of ERK 1/2 necessary to establish synaptic plasticity (SP) and gene expression-dependent CREB (). Furthermore, using electrophysiological approaches, we showed that iron is required for basal synaptic transmission and full expression of long- term potentiation (LTP), suggesting that ROS generated by iron is an important component for SP. Recent work showing that generation of ROS-dependent activation of NMDAR is required for the SP and hippocampal- dependent memoryCitation10 is in agreement with the potential role of iron in SP.
There is intensive research into the identification of other molecular targets of iron-mediated oxidation, although to date not all processes modulated by iron have been uncovered.
Given the high sensibility of DNA to iron- generated ROS,Citation11 it is intriguing to speculate about the normal function of iron in gene regulatory mechanisms. In particular, methylation of cytosine residues that are adjacent to a guanosine base is an epigenetic mechanism that has been implicated in the regulation of genes involved in SP and memory.Citation12,Citation13 Interestingly, methylation sites are frequently clustered in regulatory regions called CpG islands, which are particularly sensitive to oxidative damage.Citation14 Thus, iron- dependent oxidative reactions could also regulate gene expression by oxidizing DNA; leading to the formation of different adduct bases which may alter gene expression (). In fact, the 8hydroxyguanosine (8OHG), has been described as a marker of DNA damage.Citation11 However, the 5hydroximethylcytosine (5hmC) is formed by enzymatic oxidation over the 5methylcytosine catalyzed by a family of iron-dependent dioxygenases, recently identified in neurons.Citation15,Citation16 The functional significance of these oxidative modifications is unknown; however, they have been involved in the dynamics of DNA methylation.
Finally, combined data from our study suggest that the oxidative effect of iron is critical to activate processes that are downstream of NMDAR activation which are required for SP, like calcium release from ryanodine- sensitive intracellular store or gene expression.
Acknowledgments
We thank O. Schmachtenberg and K. Whitlock for critically reading the manuscript. This work was supported by Millennium Scientific Initiative Grant ICM, FONDAP and FONDECYT. Centro Interdisciplinario de Neurociencias de Valparaíso is a Millennium Institute.
References
- Kehrer JP. The Haber-Weiss reaction and mechanisms of toxicity. Toxicology 2000; 149:43 - 50; http://dx.doi.org/10.1016/S0300-483X(00)00231-6; PMID: 10963860
- Cheah JH, Kim SF, Hester LD, Clancy KW, Patterson SE 3rd, Papadopoulos V, et al. NMDA receptor-nitric oxide transmission mediates neuronal iron homeostasis via the GTPase Dexras1. Neuron 2006; 51:431 - 40; http://dx.doi.org/10.1016/j.neuron.2006.07.011; PMID: 16908409
- Salazar J, Mena N, Hunot S, Prigent A, Alvarez-Fischer D, Arredondo M, et al. Divalent metal transporter 1 (DMT1) contributes to neurodegeneration in animal models of Parkinson’s disease. Proc Natl Acad Sci U S A 2008; 105:18578 - 83; http://dx.doi.org/10.1073/pnas.0804373105; PMID: 19011085
- Jang SM, Kim JW, Kim CH, An JH, Kang EJ, Kim CG, et al. Control of transferrin expression by β-amyloid through the CP2 transcription factor. FEBS J 2010; 277:4054 - 65; http://dx.doi.org/10.1111/j.1742-4658.2010.07801.x; PMID: 20796026
- Gille G, Reichmann H. Iron-dependent functions of mitochondria--relation to neurodegeneration. J Neural Transm 2011; 118:349 - 59; http://dx.doi.org/10.1007/s00702-010-0503-7; PMID: 21161302
- Muñoz P, Humeres A, Elgueta C, Kirkwood A, Hidalgo C, Núñez MT. Iron mediates N-methyl-D-aspartate receptor-dependent stimulation of calcium-induced pathways and hippocampal synaptic plasticity. J Biol Chem 2011; 286:13382 - 92; http://dx.doi.org/10.1074/jbc.M110.213785; PMID: 21296883
- Adasme T, Haeger P, Paula-Lima AC, Espinoza I, Casas-Alarcón MM, Carrasco MA, et al. Involvement of ryanodine receptors in neurotrophin-induced hippocampal synaptic plasticity and spatial memory formation. Proc Natl Acad Sci U S A 2011; 108:3029 - 34; http://dx.doi.org/10.1073/pnas.1013580108; PMID: 21282625
- Aracena-Parks P, Goonasekera SA, Gilman CP, Dirksen RT, Hidalgo C, Hamilton SL. Identification of cysteines involved in S-nitrosylation, S-glutathionylation, and oxidation to disulfides in ryanodine receptor type 1. J Biol Chem 2006; 281:40354 - 68; http://dx.doi.org/10.1074/jbc.M600876200; PMID: 17071618
- Muñoz P, Zavala G, Castillo K, Aguirre P, Hidalgo C, Núñez MT. Effect of iron on the activation of the MAPK/ERK pathway in PC12 neuroblastoma cells. Biol Res 2006; 39:189 - 90; http://dx.doi.org/10.4067/S0716-97602006000100021; PMID: 16629179
- Massaad CA, Klann E. Reactive oxygen species in the regulation of synaptic plasticity and memory. Antioxid Redox Signal 2011; 14:2013 - 54; http://dx.doi.org/10.1089/ars.2010.3208; PMID: 20649473
- Valavanidis A, Vlahoyianni T, Fiotakis K. Comparative study of the formation of oxidative damage marker 8-hydroxy-2′-deoxyguanosine (8-OHdG) adduct from the nucleoside 2′-deoxyguanosine by transition metals and suspensions of particulate matter in relation to metal content and redox reactivity. Free Radic Res 2005; 39:1071 - 81; http://dx.doi.org/10.1080/10715760500188671; PMID: 16298732
- Feng J, Zhou Y, Campbell SL, Le T, Li E, Sweatt JD, et al. Dnmt1 and Dnmt3a maintain DNA methylation and regulate synaptic function in adult forebrain neurons. Nat Neurosci 2010; 13:423 - 30; http://dx.doi.org/10.1038/nn.2514; PMID: 20228804
- Levenson JM, Roth TL, Lubin FD, Miller CA, Huang IC, Desai P, et al. Evidence that DNA (cytosine-5) methyltransferase regulates synaptic plasticity in the hippocampus. J Biol Chem 2006; 281:15763 - 73; http://dx.doi.org/10.1074/jbc.M511767200; PMID: 16606618
- Valinluck V, Tsai HH, Rogstad DK, Burdzy A, Bird A, Sowers LC. Oxidative damage to methyl-CpG sequences inhibits the binding of the methyl-CpG binding domain (MBD) of methyl-CpG binding protein 2 (MeCP2). Nucleic Acids Res 2004; 32:4100 - 8; http://dx.doi.org/10.1093/nar/gkh739; PMID: 15302911
- Guo JU, Su Y, Zhong C, Ming GL, Song H. Hydroxylation of 5-methylcytosine by TET1 promotes active DNA demethylation in the adult brain. Cell 2011; 145:423 - 34; http://dx.doi.org/10.1016/j.cell.2011.03.022; PMID: 21496894
- Tahiliani M, Koh KP, Shen Y, Pastor WA, Bandukwala H, Brudno Y, et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 2009; 324:930 - 5; http://dx.doi.org/10.1126/science.1170116; PMID: 19372391