Abstract
In nature, plants have to face frequent fluctuations of intensity and spectral quality of their primary source of life - light, whose energy is needed to drive the processes of photosynthesis. A multilevel network of adaptations exists to help the plant to track and cope with fluctuations in the light environment. At the molecular level, the light harvesting antenna complex of photosystem II (LHCII), which collects the most significant part of the light energy, was found to play a central regulatory role by finely controlling the amount of energy delivered to the reaction centers. This is achieved by several mechanisms, which are summarized in this review. The fundamental features of the design of the photosynthetic antenna make photosynthetic light harvesting efficient, physiologically competent and flexible at the same time, ensuring high levels of plant survival and productivity within a wide range of light environments on our planet.
Introduction
The success of oxygenic photosynthesis in our biosphere is a great biological phenomenon that relies first of all upon the efficient design and adaptability to the changing environmental conditions of the molecular machinery of the photosynthetic apparatus. The intensity and spectral quality of light exhibits large temporal and spatial variation. For example, the daily photon flux available for plants grown in the deep shade of the tropical forest is more than 100 times less than that available for plants exposed to direct sunlight. The spectral quality of light, available for plants in the shade environment, is often different from that of the full sun light due to filtering by the canopy.Citation1 Plants are also frequently exposed to rapid and irregular changes in light intensity, which could be caused by clouds and sudden shading by other plants, occurring in natural biocenoses. And finally, diurnal changes in light quality and quantity inevitably modulate photosynthetic processes in plants on a day-to-day basis.
The dependency of the photosynthetic efficiency upon the light environment originates from the existence of a number of limiting factors, restricting the rate of light energy consumption. As a result of evolution under frequent exposure to low light fluxes, the photosynthetic machinery has evolved in such a way that only a small fraction of chlorophyll molecules are actually photosynthetically competent. They are the reaction center chlorophylls. The rest of the chlorophyll has a purely light-collecting function and is called antenna chlorophyll. This chlorophyll is specifically assembled along with carotenoids and lipids on proteins to form various light-harvesting antenna complexes, LHC's. The latter deliver excitation energy to the reaction center.
Under low light intensities (light starvation) the photosynthetic yield will depend upon the efficiency of light energy capture by antenna pigments and its delivery to the reaction centers. It also relies on the captured light energy redistribution between photosystem I and II, which operate in series to form the electron transfer chain from water to NADP. The major fundamental limitation under elevated light intensities (light saturation) arises from differences in the rates of energy absorption and transfer to the reaction centers of photosystems and subsequent electron transport. Being much slower than energy transfer, electron transport rates fulfill the fundamental thermodynamic requirement—to minimize the uphill reactions and therefore stabilize energy, which is to be used in the chain of electron/proton transfer reactions leading to NADPH and ATP synthesis. With an increase in light intensity, the reaction centers will be progressively saturated with energy (closed) leading to the reduction of the fraction of energy utilized in photosynthesis and therefore to the build up of the “unused” potentially harmful excitation energy in the photosynthetic membrane.
The Multilevel Strategies of Plant Adaptation to the Light Environment
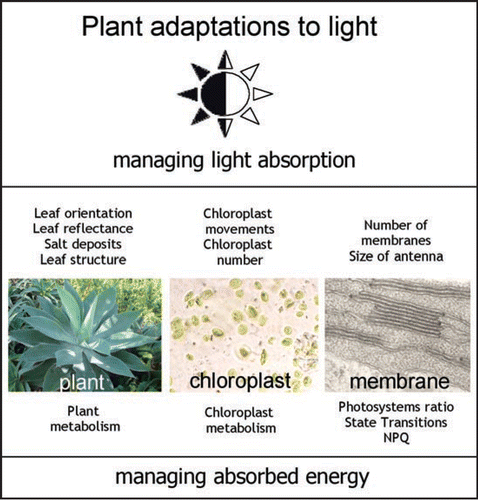
During the evolution plants have developed an entire network of adaptation mechanisms to cope with fluctuations in the light environment. These can be divided into two major groups: A—adaptations to control light absorption capacity and B—adaptations to deal with the light energy, which has been captured.
To optimize light absorption, plants respond on different levels of organization, i.e., systemically. On the level of the whole organism the adaptation involves adjustment of the leaf orientation.Citation2 This adaptation helps many land plants to cope with excess irradiation particularly during the midday. Leaf movements can be of developmental (rather slow and irreversible), passive (drought related) and active nature (reversible). The latter employs a blue-light absorbing pigment system of unknown nature as a sensor and the pulvinar motor tissue to drive leaf orientation movements.Citation3 This adaptive system is very effective in some shade plants with very low photosynthetic capacity, which are occasionally exposed to light bursts. Some desert plants have also developed a number of adaptations to increase leaf reflectance and therefore reduce the amount of absorbed light. The adaptations may include building up inorganic deposits on the leaf surface (for example, salt crystals) or developing air-filled hairs. As a rule, the efficiency of these protective methods is good but as with the developmental leaf movements, these adaptations occur on rather slow time scale.
On the cellular level, light absorption can be regulated by chloroplast movements.Citation4 These are relatively fast adaptations, occurring within minutes, but are only able to reduce light absorption by 10–20% in the environment of excess light.Citation5 The similarity of the action spectra for chloroplast and leaf movements suggests that they possess common photoreceptors.Citation5 Factors limiting chloroplast movements could be the presence of obstacles such as large vacuoles and other cellular organelles. Generally, adaptations on the cell level or at the level of the whole plant have limited capacity. In the natural environment light tends to be scattered in all directions by clouds, fog or simply high air humidity. External scattering of the incident light may substantially reduce the various leaf orientation adaptations. The absorbed light energy is also often highly scattered within the leaf by starch and various leaf tissue structures making chloroplast movements even less effective.
A most profound type of plant adaptation to light occurs at the molecular level. The type A adaptations, the regulation of light absorption, occur by long-term control of chlorophyll content in leaves. Other responses involve short-term adaptations of the photosynthetic membrane consisting of dynamic changes in light harvesting antenna size and efficiency.
Long-Term Light Adaptations: Acclimation
Acclimation is predominantly of developmental nature, and often is a result of regulation of the complex light-dependent gene expression, occurring on transcriptional, translational and post-translational levels.Citation6 This process takes days and weeks and on the level of the thylakoid membrane involves significant compositional and structural alterations. One of these is modulation of PSI and PSII antenna size by light intensity. This is a well documented and highly conserved phenomenon.Citation7 In plants, grown under high light intensity the antenna is always smaller than in those grown in shade. It was found that only the outer-LHCII complex (peripheral part of light harvesting antenna complex of photosystem II) is involved in acclimative modulation of the PSII antenna size. It was discovered that the reduction in the amount of outer-LHCII is caused by the proteolysis of its apoprotein.Citation8 The protease is suggested to be of serine and/or cysteine type of ATP-dependent enzyme, located extrinsically on the stroma side of the thylakoid membrane. It takes up to two days for the enzyme expression/post-translational activation after exposing plants to the high light environment. Once activated, it takes less than a day to complete proteolysis and reduce the outer-LHCII by a half, which is approximately 30% of the total LHCII content. “Wasting” this amount of LHCII will release about 160 kDa of the protein mass and 80 chlorophyll molecules per PSII unit—a significant metabolic event.
The other long-term acclimative responses of the thylakoid membrane to the change in the light environment (quantity and quality) include an alteration of the ratio between a number of photosystem I and II units and inactivation of a subpopulation of PSII reaction centers.Citation9,Citation10 For example, shade plants grown under light filtered on the forest canopy (more far-red light) will possess a higher PSII/PSI ratio to compensate for the reduction in the amount of red light, which is required to excite PSII.Citation1 Various aspects of the signal reception pathway, factor—receptor—transducer—gene are still not well understood. Redox state of the plastoquinone pool is suggested to play an important role in the governing multiple stress signaling and transducing mechanisms.Citation7 It has been proposed that protein phosphorylation (see below), including LHCII, plays a key part in the signal transduction path in the light acclimation processes.Citation8
Major Determinants of the Efficiency of the Photosynthetic Light Harvesting
Before introducing the short-term molecular light adaptation strategies it is important to define what determines the efficiency of the light energy utilization in the photosynthetic membrane. Two fundamental properties of the photosynthetic light harvesting antenna determine its functioning: (1) absorption cross-section—the size and spectral breadth and (2) excitation energy lifetime—the time of energy storage in the antenna before it reaches reaction centers. Cross-section depends on the type and amount of pigments, spectral redistribution of extinction coefficients and the oscillator strength. The first two factors are adjustable and determine the difference between PSI and PSII antennae. Usually PSII antenna has more pigments than that of PSI. Spectrally, PSI and PSII are also distinct. The reason for this is different absorption of their reaction center chlorophyll. Whilst the PSI reaction center absorbs at 700 nm (called P700), the PSII reaction center absorbs at 680 nm (called P680). Therefore, to match the energy of reaction center, PSI antenna absorbs longer wavelength light than PSII. This is due to enrichment in chlorophyll b in PSII, on one hand, and the presence of the long-wavelength chlorophyll a in PSI on the other. Spectral differences between PSI and PSII make the linear electron transfer efficiency potentially vulnerable to changes in the spectral quality of light and require the existence of suitable compensatory mechanisms.
The time during which antenna remains excited is the other important feature. It is generally proportional to the probability of energy delivery to the reaction center. The excitation lifetime largely depends on the chlorophyll environment. The latter could influence chlorophyll conformation, directly interfere with the optical p-electron configuration or remove excitation via energy transfer. Carotenoids, amino acids and lipids are potential modulators of the antenna chlorophyll lifetime. The excitation lifetime can be monitored by the fluorescence lifetime or yield measurements, which reveal their remarkable dynamics, reflecting the existence of control over the light harvesting process in plants.
Short-Term Molecular Adaptations to Low Light
Normally, during the short-term adaptations no changes in gene expression are involved. They occur within seconds and minutes and serve to counteract faster changes in the light environment, such as diurnal fluctuations in the light quantity and quality, sun flecks, light filtering by the canopy.Citation1 The most documented adaptation to the light quality resulting in the imbalance of the energy absorbed by photosystems is known as the State Transitions.Citation11 This process occurs within minutes and is effective at low/moderate light intensity, for example when light intensity is not limiting photosynthesis.Citation12
The molecular mechanism involved is designed to restore this energy imbalance, utilizing the redox state of the plastoquinone poolCitation13,Citation14 or/and cytochrome bCitation15 as a sensor/transducer, which are involved in activation of a protein kinase.Citation16,Citation17 When the energy imbalance is in favor of PSII, this kinase phosphorylates some polypeptides of LHCII complex, targeting Thr or Ser residues of the stroma-exposed N-terminus.Citation18 A so-called “mobile” LHCII (originating exclusively from the outer-LHCII pool) becomes dissociated from the PSII under the influence of the negative charge, introduced by phosphate.Citation19 As a consequence the PSII antenna becomes reduced. Phosphorylation leads to a partial unstacking of the grana and the detached phospho-LHCII can migrate into those regions of the thylakoid membrane, which are enriched in PSI (grana margins and stroma lamellae), and interact with the PSI complex. According to the low temperature fluorescence experiments phospho-LHCII brings to PSI 20–35% of additional excitation energy.Citation20 Assuming approximately 30% of PSII antenna decrease and about the same increase in PSI antenna, the total energy balance change between photosystems will reach 85%. This is large enough to be considered as significant for the adaptation to the changes in the light environment, particularly in shade growing plants, which have a larger pool of the peripheral LHCII.
Photosynthesis under High Light: Photoinhibition
In high light the build-up of the excess excitation energy in antenna systems will inevitably lead to photoinhibition, a sustained decline in the photosynthetic efficiency and productivity, associated with the damage of photosynthetic reaction centers.Citation21 The reaction center of the PSII is more susceptible to the damage, because of a very strong oxidation potential of the P680 (∼1,17 V) needed to oxidize water (see above). Under some conditions, when electron donation to P680 is less efficient than oxidation, an increase in the P680+ lifetime will occur. The powerful oxidant P680+ will inevitably oxidize the nearest pigments and amino acids, causing their degradation and the subsequent D1 degradation will follow.Citation22 In other circumstances, when the acceptor side is less efficient, a radical pair is formed. The recombination of this pair will lead to the P680 triplet formation. In this state P680 can interact with the atmospheric triplet oxygen, causing formation of singlet oxygen, which in turn will bleach P680.Citation23 Therefore, the number of active PSII units will be decreased and because of the slow D1 repairCitation24 the decline in electron transfer will remain even when excess light is no longer present.
Regulation of the Photosystem II Efficiency
A great deal of evidence exists that PSII efficiency can be regulated.Citation25,Citation26 It was shown that it depends reversibly upon the light intensity in a clearly non-linear manner. This contradicted the view that this dependency was associated entirely with the proportion of open reaction centers, which assumes a linear relationship between their number and the PSII quantum yield. The most feasible explanation of the observed non-linearity is opening of excess energy dissipation channel in PSII antenna. The latter became possible to detect by the chlorophyll fluorescence measurements in so-called quenching analysis using pulse-amplitude modulation technique.Citation27 One of the parameters of this analysis is called non-photochemical fluorescence quenching (NPQ). It measures the extent of the excess energy dissipation in PSII, when reaction centers are closed, i.e., not receiving excitation energy from antenna. Quantum efficiency of PSII was found to be in reciprocal linear relationship with NPQ.Citation26 This gave reasons to believe that PSII efficiency is under control of nonradiative energy dissipation, an NPQ-underlying process, which is triggered by the establishment of the proton gradient across the thylakoid membrane (ΔpH).Citation28
The Site of NPQ
For a number of years several lines of evidence suggesting that the site of NPQ is localized in LHCII antenna and not in the PSII reaction center have been acquired.Citation29 Quenching is associated with a significant decrease in the antenna fluorescence when all reaction centers are open (Fo).Citation30 The process persists if samples are frozen to 77 K and is associated with quenched fluorescence bands originating from LHCII.Citation31 Time resolved fluorescence data recorded for leaves are consistent with quenching in the antenna.Citation32 Direct measurement of heat emission in NPQ state shows it to occur within 1.4 µsec, much faster than estimates for rates of the recombination reactions in PSII.Citation33 Cross-linkers block NPQ and also transition of isolated LHCII complexes into a dissipative state.Citation34 NPQ and LHCII respond in the same way to a number of factors: antimycin A, tertiary amines and magnesium.Citation35,Citation36 NPQ is almost entirely dependent upon the presence of exclusively LHCII-bound xanthophylls, lutein and zeaxanthin.Citation36,Citation37
The Mechanism of Photoprotective Energy Dissipation
In order to understand the molecular mechanism leading to NPQ it is necessary to view it as a sequence of following events: trigger—site—change—quencher. The identities of the trigger, ΔpH, and the site, LHCII, have been introduced above. It is important to establish where exactly protons bind to the antenna and what kind of change occurs as a result of the protonation. Finally, it is necessary to identify a molecule, which act as an excitation energy trap or quencher.
Reagent DCCD, which reacts with aspartate or glutamate residues of some proteins, was found to bind to LHCII proteins and inhibit NPQ.Citation38 Two DCCD-binding amino acids have been identified on one of the LHCII complexes.Citation39 It is tempting to conclude that the proton binding directly results in the induction of quenching. DCCD binding can either inhibit the change leading to NPQ, or block the proton pathway leading to the site of quenching. Thus, the LHCII proteins have a function not just in light harvesting but also in proton translocation, and the latter function is involved in some way in NPQ.
The knowledge of the event triggering energy dissipation has been enriched by the discovery of an involvement of the protein PsbS in NPQ.Citation40 Initially thought to be a carrier of the quencher,Citation40 it is believed now to be not a pigment-binding protein.Citation41 Two lumen-exposed glutamates of PsbS have been found to bind protons.Citation42 It was also shown that ΔpH induces monomerization of the dimeric PsbS and its lateral migration out of PSII domains of the thylakoid membrane.Citation43,Citation44 Plants lacking this protein lost the ability to fast-track light intensity changesCitation45 but only marginally lost the photoprotection capacity under constant high light.Citation46 The new evidence is emerging that this protein plays a role of LHCII mobility or dynamics enhancer (Goral and Mullineaux, personal communications). Indeed being a highly hydrophobic protein PsbS could affect the fluidity of the crowded photosynthetic membraneCitation47 and therefore influence the change LHCII undergoes in order to switch into NPQ state.
The change in antenna underlying NPQ is currently in the focus of intense studies. It was proposed that protonation causes aggregation of LHCII in the membrane.Citation48 Isolated aggregated LHCII was found to be in highly dissipative state; therefore the “aggregation model” for NPQ has become common for explaining the change. According to the model, the extent of aggregation and therefore NPQ can be controlled by peripherally-bound xanthophylls, violaxanthin and zeaxanthin.Citation29 Violaxanthin, a highly polar xanthophyll, was suggested to inhibit LHCII aggregation, whereas hydrophobic zeaxanthin—to promote the process. Elevated light intensities cause conversion of violaxanthin into zeaxanthin by the enzyme de-epoxidase. In the dark another enzyme, epoxidaze, converts zeaxanthin back into violaxanthin by attaching back two epoxy oxygens to the molecule. The interconversion process of these xanthophylls was called xanthophyll cycle.Citation49 De-epoxidase is activated by ΔpH, whilst epoxidase is active when the gradient is absent. It has been proposed that the action of the xanthophylls cycle carotenoids is to allosterically control NPQ by modulating the transition of LHCII antenna in to a dissipative photoprotective state.Citation50 LHCII aggregation was a “first approximation” of this conformational change and further investigations have revealed that the dissipative state in LHCII is not a direct result of protein-protein interactions during the aggregation process but rather a consequence of intrinsic conformational change within the monomeric unit of the complex induced by protonation and/or environmental changes, such as protein aggregation.Citation34
The nature of the NPQ quencher has been addressed in several studies. The formation of chlorophyll-chlorophyll associates possessing properties of energy traps has been suggested.Citation51 However, calculations and ultrafast absorption techniques have revealed that at least the configuration of steady-state chlorophyll dimers is unlikely to be of a quenched natureCitation52 and instead, intrinsically-bound xanthophyll, lutein, has been proposed to play a role of quencher.Citation53 Indeed, carotenoids are efficient energy sinks, provided they interact very closely and specifically with chlorophyll. The atomic structure of LHCII shows that lutein molecules are indeed in a very close contact with some chlorophylls, having high dipole-dipole interaction strength.Citation54 The latter is important for the energy transfer from chlorophyll to carotenoid. In other studies, it has been found that xanthophylls cycle carotenoid, zeaxanthin can form a radical pair with chlorophyll of some minor LHCII complexes.Citation55,Citation56 This radical state was suggested to play a role in the formation of another NPQ quencher. The debates on the relative impact of lutein and zeaxanthin on NPQ are under way.Citation56,Citation57 One point is clear, though, since the mutants lacking both of these xanthophylls show no NPQ, that both of them are important pigments for the photoprotective energy dissipation in LHCII antenna.Citation37,Citation57
Concluding Remarks
Plants adapt to light in a number of different ways and on different levels of organization: whole plant, cellular and molecular (). The systemic character of adaptations simply broadens the adaptability range. Whilst the low light molecular adaptations balance the energy capture rate and distribution into two photosystems, the high light adaptations act to avoid absorption and to safely deal with the captured energy. The long-term adaptations modulate the short-term strategies. For example, there is coherence between the long-term light adaptation and the short-term adaptation to the light quality, since under limiting light the highly abundant mobile part of LHCII antenna will ensure maximum alteration in the cross-section of photosystem I and II. High light growth environment frequently leads to saturation of photosynthesis, which makes light-quality adaptation obsolete. At this point the excess light dissipation becomes a dominant adaptation. The maximum extent of NPQ at light saturation is also dependent on the plant growth conditions, mainly the type of light and temperature environment and the type of plant species. It is clear that the published values for NPQ in crop plants are relatively low.Citation58 Conversely, in plants adapted for growth in adverse environments, the extent of NPQ is three-fold higher than in the plants that have been studied most extensively in the laboratory.Citation59 Therefore it is likely that there is genetic variability in the capacity for NPQ in a manner that is related to the natural habitat of the plant. This idea has been confirmed by the screening a large number of ecologically contrasting species.Citation58
The biochemical basis of NPQ variability is either related to the composition of LHCII, including PsbS, xanthophyll cycle and ΔpH. It is known that, when plants are grown in high light, the amount of LHCII decreases, whilst PsbS increases. However, the situation is not as simple as this—mutants deficient in LHCII show reduced levels of quenching,Citation60 suggesting that it is not the total content of LHCII that is important rather the actual composition of LHCII proteins, both, trimeric and minor, which in turn shape the macrostructure of antenna system.
The knowledge, obtained in recent studies of plant responses to light is crucial for understanding the possible consequences of global warming and the associated effects on the world plant communities. The results and methodologies developing in these studies can be applied to the investigation of problems associated with biodiversity as a function of energy input into the ecosystem. Space exploration research utilizing plants on space stations and to investigate plant invasion strategies with the perspective of inhabiting other planets, should benefit unequivocally from using the knowledge of the photosynthetic mechanisms of plant adaptation to light regimes. This work is also valuable for the crop production, as it offers some important clues where and how genetic engineering could enhance plant resistance and productivity. It also raises the question of tighter links between fundamental research in plant physiology and genetic manipulation. Finally, the results of a fundamental nature and dynamics of the properties of photosynthetic antenna membrane proteins and their co-factors, chlorophylls and carotenoids are likely to be useful for the developing of technologies utilizing the energy of light.
Abbreviations
| PFD | = | photon flux density |
| PS | = | photosystem |
| LHCII | = | light harvesting II complex |
| ATP | = | adenosine triphosphate |
| DCCD | = | dicyclohexylcarbodiimide |
| NADPH | = | nicotinamide adenine dinucleotide phosphate |
| NPQ | = | nonphotochemical chlorophyll fluorescence quenching |
Figures and Tables
Figure 1 Multilevel strategies of plant adaptations to the light environment.
Acknowledgements
The author would like to acknowledge UK BBSRC and The Royal Society for financial support and Professor Conrad Mullineaux and Dr. Matthew Johnson for the critical reading of the manuscript.
References
- Anderson JM, Osmond B. Kyle DJ, Osmond B, Arntzen CJ. Sun-shade responses: Compromises between acclimation and photoinhibition. Photoinhibition 2001; Amsterdam Elsevier 1 - 38
- Bjorkman O, Powles SB. Leaf movement in the shade species Oxalis oregana L I. Response to light level and light quality. Carneg Inst Wash Yearb 1987; 80:59 - 62
- Koller D. Light-driven leaf movements. Plant Cell Environm 1990; 13:615 - 632
- Chow WS, Anderson JM, Hope AB. Variable stoichiometries of photosystem-II to photosystem-I reaction centers. Photosynth Res 1988; 17:277 - 281
- Brugnoli E, Bjorkman O. Growth of cotton under continuous salinity stress—influence on allocation pattern, stomatal and nonstomatal components of photosynthesis and dissipation of excess light energy. Planta 1992; 187:335 - 347
- Kloppstech K. Light regulation of photosynthetic genes. Physiol Plant 1997; 100:739 - 747
- Anderson JM, Chow WS, Park YI. The grand design of photosynthesis: Acclimation of the photosynthetic apparatus to environmental cues. Photosynth Res 1995; 46:129 - 139
- Andersson B, Aro EM. Proteolytic activities and proteases of plant chloroplasts. Physiol Plant 1997; 100:780 - 793
- Anderson JM, Chow WS, Goodchild DJ. Thylakoid membrane organisation in sun/shade acclimation. Aust J Plant Physiol 1988; 15:11 - 26
- Melis A. Dynamics of photosynthetic membrane-composition and function. Biochim Biophys Acta 1991; 1058:87 - 106
- Bonaventura C, Mayers J. Fluorescence and oxygen evolution from Chlorella pyrenoidosa. Biochim Biophys Acta 1969; 189:366 - 383
- Horton P, Hague A. Studies on the induction of chlorophyll fluorescence in isolated barley protoplasts 4. Resolution of non-photochemical quenching. Biochim Biophys Acta 1988; 932:107 - 115
- Horton P. The effect of redox potential on the kinetics of fluorescence induction in peachloroplasts 2. Sigmoidicity. Biochim Biophys Acta 1981; 637:152 - 158
- Allen JF, Bennett J, Steinback KE, Arntzen CJ. Chloroplast protein phosphorylation couples plastoquinone redox state to distribution of excitation energy between photosystems. Nature 1981; 291:1 - 5
- Gal A, Shahak Y, Schuster G, Ohad I. Specific loss of LHCII phosphorylation in the Lemna mutant-1073 lacking the cytochrome-b6/f complex. FEBS Letts 1987; 221:205 - 210
- Bennett J. Phosphorylation of chloroplast membrane proteins. Nature 1977; 269:344 - 346
- Bennett J. Chloroplast phosphoproteins. The protein kinase of thylakoid membranes is light-dependent. FEBS Letts 1979; 103:342 - 344
- Mullet JE. The amino-acid-sequence of the polypeptide segment which regulates membrane adhesion (grana stacking) in chloroplasts. J Biol Chem 1983; 258:9941 - 9948
- Barber J. Properties and organization of photosynthetic pigments. Symp Soc Exp Biol 1983; 36:19 - 52
- Kyle DJ, Arntzen CJ. Thylakoid membrane protein phosphorylation selectively alters the local membrane surface charges near the primary acceptor of photosystem II. Photobiochem Photobiophys 1983; 5:11 - 25
- Powles SB. Photoinhibition of photosynthesis induced by visible-light. Ann Rev Plant Physiol Plant Molec Biol 1984; 35:15 - 44
- Barber J. Molecular-basis of the vulnerability of photosystem-II to damage by light. Aust J Plant Physiol 1995; 22:201 - 208
- Telfer A, He WZ, Barber J. Spectral resolution of more than one chlorophyll electron-donor in the isolated photosystem-II reaction center complex. Biochim Biophys Acta 1990; 1017:143 - 151
- Ohad I, Kyle DJ, Arntzen CJ. Membrane-protein damage and repair—removal and replacement of inactivated 32-kilodalton polypeptides in chloroplast membranes. J Cell Biol 1984; 99:481 - 485
- Weis E, Berry JA. Quantum efficiency of Photosystem-II in relation to energy-dependent quenching of chlorophyll fluorescence. Biochim Biophys Acta 1987; 894:198 - 208
- Genty B, Briantais JM, Baker NR. The relationship between the quantum yield of photosynthetic electron-transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 1989; 990:87 - 92
- Schreiber U. Detection of rapid induction kinetics with a new type of high-frequency modulated chlorophyll fluorometer. Photosynth Res 1986; 9:261 - 272
- Oxborough K, Horton P. A study of the regulation and function of energy-dependent quenching in pea-chloroplasts. Biochim Biophys Acta 1988; 934:135 - 143
- Horton P, Ruban AV, Walters RG. Regulation of light harvesting in green plants. Annu Rev Plant Physiol Plant Mol Biol 1996; 47:665 - 684
- Horton P, Ruban AV. Delta-pH-dependent quenching of the Fo-level of chlorophyll fluorescence in spinach leaves. Biochim Biophys Acta 1993; 1142:203 - 206
- Ruban AV, Rees D, Noctor GD, Young A, Horton P. Long-wavelength chlorophyll species are associated with amplification of high-energy-state excitation quenching in higher-plants. Biochim Biophys Acta 1991; 1059:355 - 360
- Genty B, Goulas Y, Dimon B, Peltier G, Briantais JM, Moya I. Modulation of efficiency of primary conversion in leaves. Photosynth Res 1992; 34:106
- Mullineaux CW, Ruban AV, Horton P. Prompt heat release associated with delta-pH-dependent quenching in spinach thylakoid membranes. Biochim Biophys Acta 1994; 1185:119 - 123
- Ilioaia C, Johnson M, Horton P, Ruban AV. Induction of efficient energy dissipation in the isolated light harvesting complex of photosystem II in the absence of protein aggregation. J Biol Chem 2008; 283:29505 - 29512
- Noctor G, Ruban AV, Horton P. Modulation of delta-pH-dependent nonphotochemical quenching of chlorophyll fluorescence in spinach-chloroplasts. Biochim Biophys Acta 1993; 1183:339 - 344
- Ruban AV, Young A, Horton P. Modulation of chlorophyll fluorescence quenching in isolated light-harvesting complex of photosystem-II. Biochim Biophys Acta 1994; 1186:3 - 127
- Niyogi KK, Shih C, Chow WS, Pogson BJ, DellaPenna D, Björkman O. Photoprotection in a zeaxanthin and lutein deficient double mutant of Arabidopsis. Photosynth Res 2001; 67:139 - 145
- Ruban AV, Walters RG, Horton P. The molecular mechanism of the control of excitation-energy dissipation in chloroplast membranes—inhibition of delta-pH-dependent quenching of chlorophyll fluorescence by dicyclohexylcarbodiimide. FEBS Letts 1992; 309:175 - 179
- Walters RG, Ruban AV, Horton P. Identification of proton-active residues in a higher plant light-harvesting complex. Proc Nat Acad Sci USA 93:14204 - 14209
- Li X-P, Bjorkman O, Shih C, Grossman AR, Rosenquist M, Jansson S, Niyogi KK. A pigment-binding protein essential for regulation of photosynthetic light harvesting. Nature 2000; 403:391 - 395
- Bonente G, Howes BD, Caffarri S, Smulevich G, Bassi R. Interactions between the photosystem II subunit PsbS and xanthophylls studied in vivo and in vitro. J Biol Chem 2008; 283:8434 - 8445
- Li XP, Gilmore AM, Caffari S, Bassi R, Golan T, Kramer D, Niyogi KK. Regulation of light harvesting involves intrathylakoid lumen pH sensing by the PsbS protein. J Biol Chem 2004; 279:22866 - 22874
- Bergantino E, Segalia A, Brunetta A, Teardo E, Rogoni F, Giacometti GM, Szabo I. Light- and pH-dependent structural changes in the PsbS protein of hotosystem II. Proc Nat Acad Sci USA 2003; 100:15265 - 15270
- Teardo E, Polverino de Laureto P, Bergantino E, Dalla Vecchia F, Rigoni F, Szabò I, Giacometti GM. Evidences for interaction of PsbS with photosynthetic complexes in maize thylakoids. Biochim Biophys Acta 2007; 1767:703 - 711
- Kiss A, Crouchman S, Ruban AV, Horton P. The PsbS protein controls the organisation of the photosystem II antenna in higher plant thylakoid membranes. J Biol Chem 2008; 283:3972 - 3978
- Li XP, Muller-Moule P, Gilmore AM, Niyogi KK. PsbS-dependent enhancement of feedback de-excitation protects photosystem II from photoinhibition. Proc Nat Acad Sci USA 2002; 99:15222 - 15227
- Kirchhoff H. Molecular crowding and order in photosynthetic membranes. Trends Plant Sci 2008; 13:201 - 207
- Horton P, Ruban AV, Rees D, Noctor G, Pascal AA, Young A. Control of the light harvesting function of chloroplast membranes by aggregation of the LHCII chlorophyll protein complex. FEBS Letts 1991; 292:1 - 4
- Yamamoto HY, Nakayama TOM, Chichester CO. Studies on the light and dark interconversions of leaf xanthophylls. Arch Biochem Biophys 1962; 97:168 - 173
- Horton P, Ruban AV, Wentworth M. Allosteric regulation of the light harvesting system of photosystem II. Phil Trans Roy Soc London B 2000; 355:1361 - 1370
- Horton P, Wentworth M, Ruban A. Control of the light harvesting function of chloroplast membranes: the LHCII-aggregation model for non-photochemical quenching II. FEBS Letts 2005; 579:4201 - 4206
- Duffy CDP, Ruban A, Barford W. Possible role of strongly-coupled chlorophyll dimers in photoprotection of LHCII. J Phys Chem 2008; 112:12508 - 12515
- Ruban AV, Berera R, Ilioaia C, Stokkum IHM, Kennis JTM, Pascal AA, van Amerongen H, Robert B, Horton P, van Grondelle R. Identification of a mechanism of photoprotective energy dissipation in higher plants. Nature 2007; 450:575 - 578
- Liu ZF, Yan HC, Wang KB, Kuang TY, Zhang JP, Gui LL, An XM, Chang WR. Crystal structure of spinach major-light harvesting complex at 2.72 Å resolution. Nature 2004; 428:287 - 292
- Holt NE, Zigmantas D, Valkunas L, Li XP, Niyogi KK, Fleming GR. Carotenoid cation formation and the regulation of photosynthetic light harvesting. Science 2005; 307:433 - 436
- Ahn TK, Avenson T, Ballottari M, Cheng YC, Niyogi KK, Bassi R, Fleming GR. Architecture of a charge-transfer state regulating light harvesting in a plant antenna protein. Science 2008; 320:794 - 797
- Johnson MP, Pérez-Bueno ML, Zia A, Horton P, Ruban AV. The zeaxanthin-independent and zeaxanthin-dependent qE components of non-photochemical quenching involve common conformational changes within the Photosystem II antenna in Arabidopsis thaliana. Plant Physiol 2008; http://dx.doi.org/10.1104/pp.109.129957
- Johnson GN, Young AJ, Scholes JD, Horton P. The dissipation of excess excitation energy in British plant species. Plant Cell Environ 1993; 16:673 - 679
- Ruban AV, Young AJ, Horton P. Induction of non-photochemical energy-dissipation and absorbency changes in leaves—evidence for changes in the state of the light-harvesting system of Photosystem-II in-vivo. Plant Physiol 1993; 102:741 - 750
- Horton P, Johnson M, Perez M, Ruban AV. Does the structure and macro-organization of photosystem II in higher plant grana membranes regulate light harvesting states?. FEBS J 2008; 275:1069 - 1079