
Abstract
Efficient assembly of a mitotic spindle and stable attachment of microtubules (k-fibers) to kinetochores are essential for the high fidelity of chromosome segregation. Both spindle assembly and Mitotic spindle mediates the segregation of chromosomes in the cell cycle and the proper function of the spindle is crucial to the high fidelity of chromosome segregation and to the stability of the genome. Nucleation of microtubules (MTs) from centrosomes and chromatin represents two well-characterized pathways essential for the assembly of a dynamic spindle in mitosis. Recently, we identified a third MT nucleation pathway, in which existing MTs in the spindle acts as a template to promote the nucleation and polymerization of MTs, thereby efficiently amplifying MTs in the spindle. We will review here our current understanding on the molecular mechanism, the physiological function and the cell-cycle regulation of MT amplification.k-fiber formation require robust nucleation and polymerization of microtubules mediated by the γ-tubulin ring complex (γ TuRC). It has been well established that centrosomes and chromatin are the two centers for microtubule nucleation. We recently demonstrate a third mechanism for microtubule nucleation and polymerization, in which the existing microtubules in the spindle act as templates to promote the formation of new microtubules. We showed that a novel spindle-associated protein, FAM29A, plays a critical role in this microtubule-dependent microtubule amplification. FAM29A associates with spindle microtubules and directly interacts with and recruits NEDD1, the targeting subunit of γTuRC. Spindle-associated γTuRC then promotes microtubule nucleation required for spindle assembly and k-fiber formation. This novel microtubule amplification pathway provides a powerful mechanism to control the local cytoskeleton structures independent of centrosomes and chromatin. We speculate that microtubule amplification not only functions in mitosis, but may also act in other physiological processes to re-enforce existing cytoskeleton structures.
Figures and Tables
Figure 1 FAM29A mediates MT amplification in spindle assembly and in k-fiber maturation. Active kinase of Plk1 targets FAM29A to spindle microtubules and FAM29A directly interacts with and recruits NEDD1, the targeting subunit of γTuRC. Spindle-associated γTuRC then promotes microtubule nucleation required for spindle assembly and k-fiber formation. Drawings for various cellular structures are representative, but not to the scale.
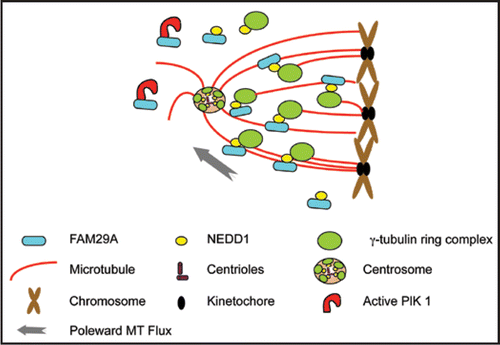
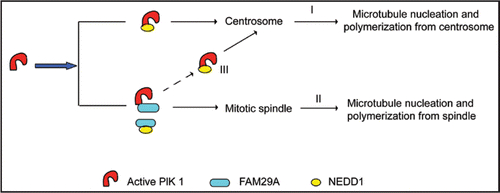
Figure 2 FAM29A controls the partition of NEDD1 between spindle MTs and centrosomes. The interaction between Plk1 and NEDD1 is responsible for Plk1-mediated recruitment of NEDD1 to centrosomes, which controls microtubule nucleation and polymerization from centrosomes (I). Recruitment of NEDD1 to the spindle depends on the interaction between Plk1 and FAM29A as well as the interaction between FAM29A and NEDD1, which mediate microtubule nucleation and polymerization from the mitotic spindle (II). The levels of FAM29A determine the partition of NEDD1 between centrosomes and the spindle, which, in turn, controls the relative contributions of MT nucleation and polymerization between these two mitotic structures (III).
Addendum to: