
Abstract
The second messenger cGMP controls cardiovascular and gastrointestinal homeostasis in mammals. However, its physiological relevance in the nervous system is poorly understood.1 Now, we have reported that the cGMP-dependent protein kinase type I (PRKG1) is implicated in the regulation of the timing and quality of sleep and wakefulness.2 Prkg1 mutant mice showed altered distribution of sleep and wakefulness as well as reduction in rapid-eye-movement sleep (REMS) duration and in non-REMS consolidation. Furthermore, the ability to sustain waking episodes was compromised. These observations were also reflected in wheel-running and drinking activity. A decrease in electroencephalogram power in the delta frequency range (1–4 Hz) under baseline conditions was observed, which was normalized after sleep deprivation. Together with the finding that circadian clock amplitude is reduced in Prkg1 mutants these results indicate a decrease of the wake-promoting output of the circadian system affecting sleep. Because quality of sleep might affect learning we tested Prkg1 mutants in several learning tasks and find normal spatial learning but impaired object recognition memory in these animals. Our findings indicate that Prkg1 impinges on circadian rhythms, sleep and distinct aspects of learning.
Acknowledgements
We like to thank Rüdiger Klein for the Nes-Cre mice, and Anne-Marie Schönegge and Claudia Becker for help with behavioral testing. This research was supported by the DFG and VolkswagenStiftung (Robert Feil), the National Genome Research Network (NGFN), Foerderkennzeichen 01GR0430, and by EUMODIC, LSHG-CT-2006-037188 (Sabine M. Hölter and Wolfgang Wurst), the Dr. Karl Kuhn-Stiftung and fortüne-Programm der Medizinischen Fakultät der Universität Tübingen (grant #1774-0-0) (Susanne Feil), and the Swiss National Science Foundation, the State of Fribourg and EUCLOCK (Urs Albrecht).
Figures and Tables
Figure 1 Light induction of Per1, Per2 and cFos in the SCN of Prkg1BKO mice (black bars, genotype: Prkg1L-/L2; Nes-Cretg/0) compared to litter-matched control mice (grey bars, genotype: Prkg1+/L2; Nes-Cretg/0). Animals were kept in a 12 h light-12 h dark cycle with water and food ad libitum. A light pulse of 500 lux intensity and 15-min duration was applied at circadian time (CT) 14 or 22. One hour after the light pulse animals were sacrificed and analyzed for gene expression. Control animals receiving no light pulse were also analyzed for gene expression (white bars). Shown are the expression levels of (A) Per1 mRNA, (B) Per2 mRNA and (C) cFos mRNA in the SCN as detected by RNA in situ hybridization. The data shown represent relative optical densities as determined by the difference of staining in the SCN relative to surrounding tissue not expressing the genes. Values are the means ± SEM of three independent experiments. Significance was determined by student's t-test (*p < 0.05, n = 3). Animals studied were males between 2–4 months of age. All experiments were in accordance with Swiss animal protection law in the declaration of Helsinki.
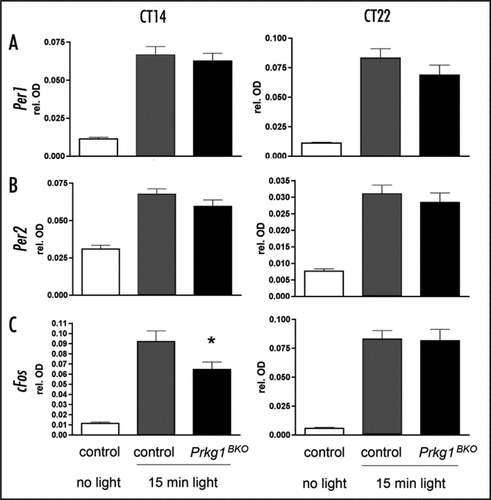
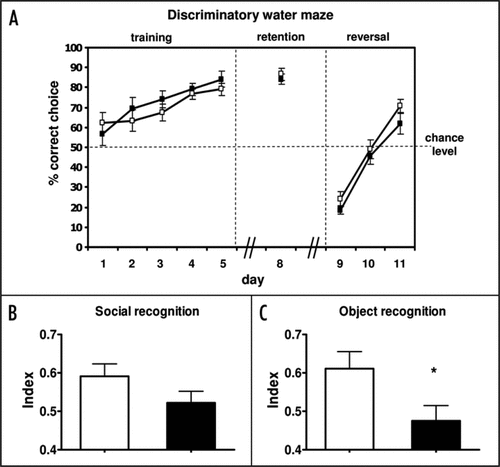
Figure 2 Analysis of cognitive functions of Prkg1BKO mice (black symbols and bars, genotype: Prkg1L-/L2; Nes-Cretg/0) compared to litter-matched control mice (white symbols and bars, genotype: Prkg1+/L2; Nes-Cretg/0). Mice were group-housed with food and water ad libitum, and maintained on a 12 h light-12 h dark cycle. Behavioral tests were performed during the light phase by an observer that was blinded to the genotype of the animals. All experiments were in accordance with German animal protection law. (A) Spatial learning was assessed in a discriminatory water maze task as described.Citation17 Eight- to twelve-month-old female mice (n = 12 per genotype) had to discriminate between two visible platforms: a stable platform remaining in the same position (correct choice) and a platform that was moved in a pseudorandom fashion and submerged when climbed by a mouse. Mice were tested over five training sessions (day 1–5) and again at day 8 (memory retention). To validate spatial learning strategies, the correct platform was then moved to the opposite quadrant (day 9–11, reversal). The number of correct choices is expressed as the percentage of the total number of choices per session (10 trials per session). The escape latencies, i.e., the times required for navigating to one of the two platforms, were not different between genotypes (data not shown). (B) Social memory was tested using the social discrimination paradigmCitation32 in 8- to 9-month-old female mice (n = 22 controls; n = 17 mutants). The procedure consisted of two 4-min exposures of stimulus animals (4-week-old male C56BL/6J mice) to the test animal in a fresh cage to which the test animal had been moved 2 h prior to testing. During the first exposure, one stimulus animal was exposed to the test animal. After a retention interval of 2 h, this stimulus animal was re-exposed to the test animal together with an additional, previously not presented stimulus animal. The duration of investigatory behavior of the test animal towards the stimulus animals was recorded by a trained observer with a hand-held computer. A social recognition index was calculated as time spent investigating the unfamiliar stimulus mouse/time spent investigating both the familiar and unfamiliar stimulus mouse. (C) Object memory was assessed in an object recognition task according to the procedure described by Genoux and co-workersCitation33 using 15-month-old female mice (n = 12 per genotype). Briefly, the mouse was allowed to explore two identical objects three times for 5 min, with an inter-trial interval of 15 min. After a retention interval of 2 h, one of the previously encountered familiar objects was substituted by a new, unfamiliar one, which was again substituted by a novel object after a second retention interval of 24 h. For each retention test the mouse was put again into the test box for 5 min, and exploration time—defined as touching the object with the nose—was recorded by a trained observer with a hand-held computer. An object recognition index was calculated as time spent investigating the unfamiliar object/time spent investigating both the familiar and unfamiliar object. The diagram shows the object recognition index of control and Prkg1BKO mutant mice after a retention interval of 24 h. Note that in this experiment after a retention interval of 2 h the index was not significantly different between genotypes (data not shown). Data obtained in the social and object recognition experiments were analyzed by using the Observer 4.1 Software (Noldus, Wageningen). All data are expressed as mean ± SEM. *p < 0.05 (one-tailed t-test).
Addendum to: