
Abstract
For many familiar with metazoan relationships and body plans, the hypothesis of a sister group relationship between Diploblasta and Bilateria1 comes as a surprise. One of the consequences of this hypothesis - the independent evolution of a nervous system in Coelenterata and Bilateria - seems highly unlikely to many. However, to a small number of scientists working on Metazoa, the parallel evolution of the nervous system is not surprising at all and rather a confirmation of old morphological and new genetic knowledge.2-4 The controversial hypothesis that the Diploblasta and Bilateria are sister taxa is, therefore, tantamount to reconciling the parallel evolution of the nervous system in Coelenterata and Bilateria. In this addendum to Schierwater et al.,1 we discuss two aspects critical to the controversy. First we discuss the strength of the inference of the proposed sister relationship of Diploblasta and Bilateria and second we discuss the implications for the evolution of nerve cells and nervous systems.
Figures and Tables
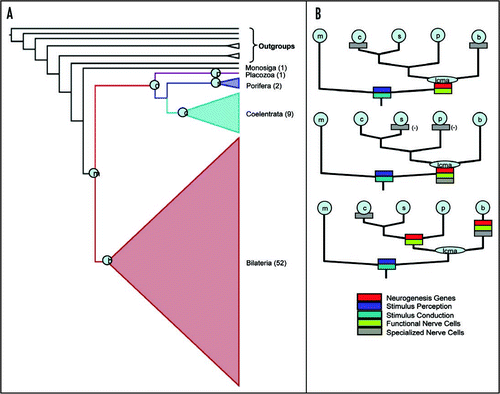
Figure 1 (A) Phyologenetic tree with relationships within Bilateria, Coelenterata and Porifera collapsed. The 72 taxa are comprised of the 64 taxa from (5) plus eight taxa added from (1). Numbers in parentheses refer to number of species in each of these groups. Phylogenetic matrices and tree topologies within the collapsed groups are available from the authors. We inferred the phylogeny using a maximum likelihood (ML) and maximum parsimony (MP) optimality criterion. Node support values (ML/MP) for nodes marked by circles with inset letters are: (B) Bilateria 100/100, (C) Coelenterata 100/82, (S) Porifera 100/100, (D) Diploblasta 100/99, (M) Metazoa 100/63; (P) Placozoa is a single taxon. Within the Bilateria: Deuterostomia 100/100, Protostomia 100/100. (B) Phylogenetic scenarios for the evolution of nerve cells mapped onto the Diploblast-Bilateria Sister hypothesis. Five potential characters (represented by colored boxes in the figure) important in the evolution of nerve cells are mapped onto the Diploblast-Bilateria Sister. Most qualities of a nerve cell seem to have been present already in the last common metazoan ancestor (LCMA in light blue). In the top figure we present the most parsimonious explanation for the evolution of these five characters (6 parsimony steps). Only the specialization of multifunctional proto-nerve cells into unifunctional nerve cells would have occurred in parallel in Bilateria and Coelenterata in the above scenario. The middle scenario is similar to the top only instead of hypothesizing independent gain of specialized nerve cells it hypothesizes independent loss of specialized nerve cells (7 steps). The bottom tree shows a highly unlikely scenario where the number of steps is nearly twice that of the top scenario.
Addendum to: