
Abstract
Nociceptive sensitization is a conserved form of neuronal plasticity that serves an important survival function, as it fosters behavior that protects damaged tissue during healing. This sensitization may involve a lowering of the nociceptive threshold (allodynia) or an increased response to normally noxious stimuli (hyperalgesia). Although nociceptive sensitization has been intensively studied in vertebrate models, an open question in the field is the extent to which allodynia and hyperalgesia, which almost always occur in tandem, are truly separate events at the mechanistic level. We recently introduced a genetically tractable model for damage-induced nociceptive sensitization in Drosophila larvae, and identified a conserved cytokine signaling module that mediates development of allodynia following UV irradiation. This pathway includes the Drosophila homolog of Tumor Necrosis Factor-alpha (TNFα), Eiger, which is released from damaged epidermal cells and acts directly on its receptor, Wengen, located on nociceptive sensory neurons. Here we show that although Eiger and Wengen are both required for the development of thermal allodynia, they are dispensable for thermal hyperalgesia, suggesting, contrary to what is commonly assumed, that these two forms of hypersensitivity are initiated by separate genetic pathways.
Acknowledgements
This work was supported by an American Heart Association predoctoral fellowship (0815339F) to DTB and University of Texas MD Anderson institutional startup funds to MJG. The authors declare that they have no competing financial interests.
Figures and Tables
Figure 1 Timing of UV-induced responses. Both allodynia and hyperalgesia develop before overt morphological evidence of epidermal damage and cleaved caspase immunoreactivity indicative of programmed cell death. Although mild allodynia arises earlier than hyperalgesia, the peak response and subsequent diminishment of hyperalgesia occur more rapidly. Staining using a stress response reporter (msn-lacZ, which reads out activation of the Jun N-Terminal Kinase signaling pathway) was evident from 4 to 48 hours after UV treatment. Solid lines, times response was seen most often. Dashed lines, times response was occasionally observed, or was mild or diminishing.
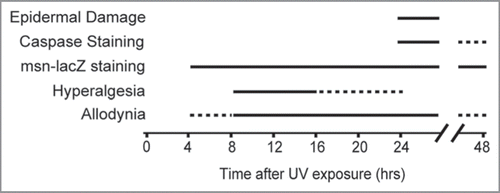
Figure 2 Eiger and Wengen do not affect the development of thermal hyperalgesia. Whole-animal eiger null mutants (egr1/egr3) or tissue-specific expression of RNAi transgenes targeting eiger (UASeigerIR) or wengen (UAS-wengenIR) were used to test development of thermal hyperalgesia 8 hours following UV irradiation. Epidermal-specific (via A58-Gal4) knockdown of Eiger and nociceptor-specific (via ppk-Gal4) knockdown of Wengen do not prevent thermal hyperalgesia following UV damage. Withdrawal latency was measured (in seconds) in response to a thermal probe heated to 45°C. Larvae of all genotypes treated with UV (gray bars) respond nearly twice as fast as their mock-treated counterparts (black bars). Error bars = SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
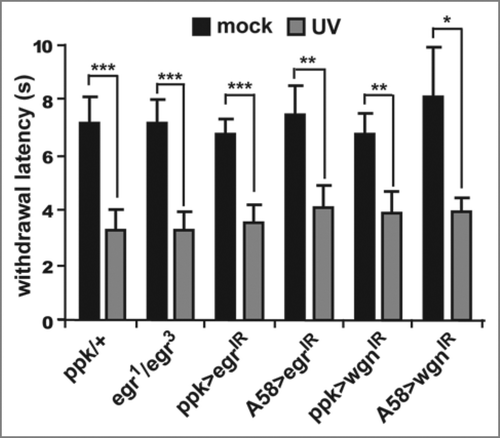
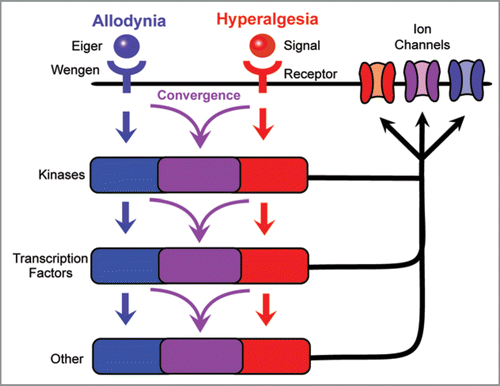
Figure 3 Possible relationships of signaling cascades mediating thermal allodynia and hyperalgesia. The development of thermal allodynia and thermal hyperalgesia is regulated by distinct receptors and may involve completely separate signaling pathways with minimal to no overlap. By contrast, these signals could converge downstream of receptor activation, allowing a single component to regulate multiple aspects of nociceptive sensitization. This convergence may take place at the level of cytoplasmic kinases, nuclear transcription factors, or other intracellular mediators, or the ion channels that are the likely endpoints of each signaling pathway.
Addendum to: