Abstract
From a structural standpoint, living organisms are organized like a nest of Russian matryoshka dolls, in which structures are buried within one another. From a temporal point of view, this type of organisation is the result of a history comprised of a set of time backcloths which have accompanied the passage of living matter from its origins up to the present day. The aim of the present paper is to indicate a possible course of this passage through time and suggest how today’s complexity has been reached by living organisms. This investigation will employ three conceptual tools, namely the Mosaic, Self-Similarity Logic, and the Biological Attraction principles. Self-Similarity Logic indicates the self-consistency by which elements of a living system interact, irrespective of the spatio-temporal level under consideration. The term Mosaic indicates how, from the same set of elements assembled according to different patterns, it is possible to arrive at completely different constructions: hence, each system becomes endowed with different emergent properties. The Biological Attraction principle states that there is an inherent drive for association and merging of compatible elements at all levels of biological complexity. By analogy with the gravitation law in physics, biological attraction is based on the evidence that each living organism creates attractive ‘field’ around itself. This field acts as a sphere of influence that is actively attracting similar fields of other biological systems, thereby modifying salient features of interacting organisms. Three specific organizational levels of living matter, namely the molecular, cellular, and supracellular levels, have been considered in order to analyse and illustrate the interpretative as well as the predictive roles of each of these three explanatory principles.
Note
This article is dedicated to Gianpiero Turrin, great friend and man of great culture.
Figures and Tables
Figure 1 Schematic representation of the well of the past.
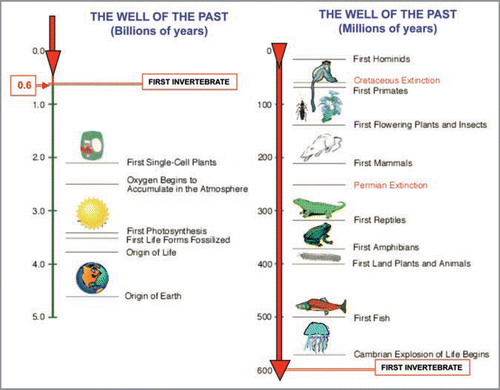
Figure 2 Schematic representation of the ‘Russian Doll Principle’ proposed by Jacob.

Figure 3 Schematic representation of miniaturization and hierarchic criteria. The four main miniaturization levels, i.e., the macro-, meso-, micro- and nano-scales are illustrated. Furthermore, mechanisms for the plastic adjustment of the 3D-conformations of proteins are outlined in view of their functional importance since as stated by JacobCitation6: “the overall coordination of the cellular function is founded on the geometrical deformation of proteins”.Citation6
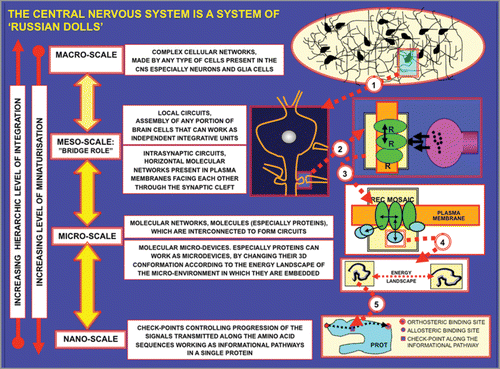
Figure 4 Schematic representation of the ‘Mosaic Principle’. The left panel shows the mosaic in Ravenna of Empress Teodora made up by small cubes of coloured marble (tesserae). As the right panel shows, from the same set of protein monomers (tesserae) it is possible to assemble completely different multimeric proteins.
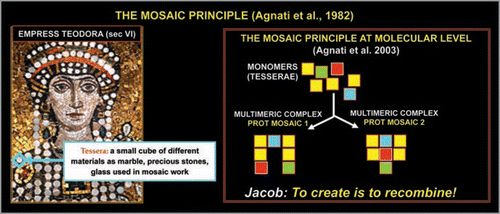
Figure 5 Schematic representation of a multidomain protein.
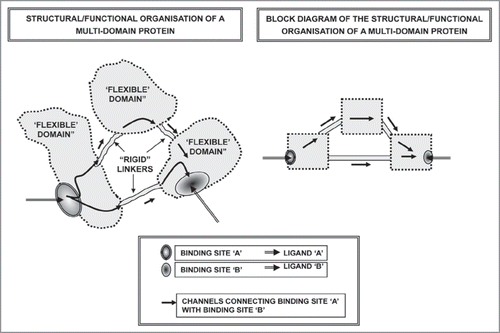
Figure 6 Schematic representation of the possible basic mechanisms involved in the assemblage of multimeric proteins.
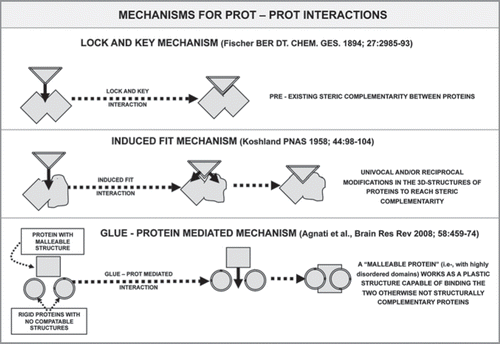
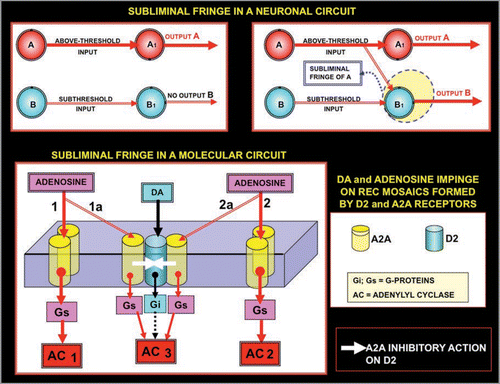
Figure 7 Schematic representation of the model of lateral inhibition transferred according to the ‘Self-Similarity Logic’ principle: from neural networks to molecular networks. Upper panel: Lateral inhibition leads to focusing of activity in a few computational neurons in neural circuits.Citation39 Lower panel: The scheme illustrates the possible application of this basic model of circuit organization to a complex formed by a A2A^D2 heterodimers and A2A homodimers. Activation of A2A receptors by Adenosine leads to a reduction in the affinity of D2 receptors in the heterodimers, hence in a sharpening of the signalling via the A2A homodimers. For further details, see text.
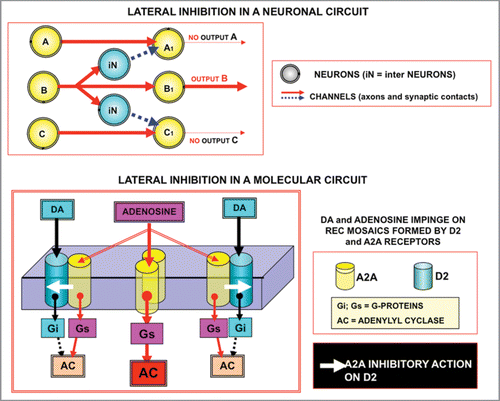
Figure 8 Schematic representation of the concept of fringe. Two input neurons, A and B, directly acting on two output neurons (A1 and B1) are considered. It is surmised that A, but not B (upper left panel), gives an above-threshold input to its target neuron A1 (upper left panel). However, if neuron A fires at a high frequency it can lower the threshold for the firing of B1. The term ‘mosaic’ will be used to describe not only the physical contact and assemblage of elements, but also (as mentioned above) at a later more developed organimal complexity when individual building blocks become functionally inter-related and inter-dependent, as is the case of cellular organelles within the cytoplasm. Thus, the subthreshold input of B to B1 is now sufficient to cause the firing of neuron B1, hence two output neurons fire (upper right panel). Thus, these panels show “facilitation” of A on the firing of neuron B1, and indicate that B1 is localised in the subliminal fringe of A. The same basic scheme is applied (lower panel) to receptor mosaics formed by A2A and D2 receptors. A low concentration of Adenosine can activate only AC1 and AC2, however high concentrations of Adenosine can diffuse until the trimer A2A^D2^A2A inhibiting, via receptor-receptor interactions, the activation by D2 of the Gi. Thus, it can be observed that the activation is not simply of AC1 and AC2, but also of AC3, which is no longer inhibited by the Gi