Abstract
The urgent requirement of next generation antimalarials has been of recent interest due to the emergence of drug-resistant parasite. The genome-wide analysis of Plasmodium falciparum helicases revealed three RuvB proteins. Due to the presence of helicase motif I and II in PfRuvBs, there is a high probability that they contain ATPase and possibly helicase activity. The Plasmodium database has homologs of several key proteins that interact with RuvBs and are most likely involved in the cell cycle progression, chromatin remodeling, and other cellular activities. Phylogenetically PfRuvBs are closely related to Saccharomyces cerevisiae RuvB, which is essential for cell cycle progression and survival of yeast. Thus PfRuvBs can serve as potential drug target if they show an essential role in the survival of parasite.
Current Scenario of Understanding of RuvB Family of Enzymes
The availability of complete genome sequence of Plasmodium falciparum at Plasmodb (www.PlasmoDB.org) makes feasible the study of the essential genes important for parasite biology and pathogenesis. The parasite specific genes important for various pathways can serve as good targets for antimalarial drug. The genome-wide analysis of P. falciparum helicases suggested that parasite specific helicases can be good antimalarial drug targets.Citation1 P. falciparum RuvB (PfRuvB) proteins are structural homologs of bacterial RuvB protein. Two homologs of RuvB in Saccharomyces cerevisiae (scRuvBL1 and scRuvBL2) have been studied in detail and the knockout of scRuvBL1 demonstrated its essentiality in yeast growth.Citation2 Previously it has been reported that RuvB either itself or with the association of some interacting partner is involved in the holliday junction formation, branch migration and resolution of holliday junction in prokaryotes. It has also been well established that RuvBs are involved in the replication fork reversal both in prokaryotes and eukaryotes.
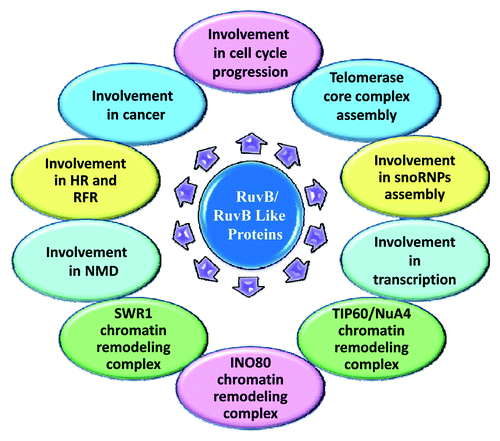
Further studies in yeast and some other systems demonstrated that RuvBs are involved in multiple cellular pathways (), such as cell cycle progression and RNA polymerase II-directed transcription,Citation3 DNA damage response, replication fork reversal,Citation4 nonsense-mediated mRNA decay,Citation5 insulin stimulated GLUT4 translocation,Citation6 small nucleolar ribonucleotide protein (snoRNPs) assembly,Citation7,Citation8 cellular transformation,Citation9 cancer metastasis, apoptosis, mitosis, and development.Citation10 RuvBs are essential components of several multiprotein complexes () and it is expected that their mode of function was very similar in these wide range of complexes.Citation8 Considering all the above roles and involvement in several important cellular pathways in different systems, P. falciparum RuvB proteins are very similar to those of S. cerevisiae essential RuvB like Protein and can be expected to play essential role in the parasite cell cycle progression and chromatin remodeling. Therefore it has been of recent interest to explore the RuvB family of proteins from P. falciparum in detail and study their role in various activities in the malaria parasite. Emergence of drug-resistant P. falciparum parasite severely affects the prevention and treatment of malaria (Box 1). Thus it has been of current interest to identify novel drug target and new antimarials to fight the drug-resistant parasite. Recent work has shown that RuvBs have some role in cancer. A recent report showed that Reptin (RuvBL2) is required for the transcription of telomerase reverse transcriptase and is overexpressed in gastric cancer.Citation15 In another report, the in vivo silencing of Reptin (RuvB2) resulted in the blockage of human hepatocellular carcinoma and was associated with replicative senescence.Citation16 Here in this review we will focus on RuvB proteins of P. falciparum, along with its homologs in yeast, E. coli, human and other systems.
Figure 1. Schematic diagram of RuvB/RuvB like proteins involvement in diverse cellular activities. This schematic of RuvB proteins involved in various activities was prepared on the basis of reported role in E. coli, yeast, drosophila and human etc. RuvB/RuvB like proteins are involved in the diverse cellular activities shown in the figure.
RuvB Family of Enzymes Belong to AAA+ Class Enzyme
RuvB belongs to a category of proteins called the AAA+ class (ATPases associated with various cellular activities), which contains one or two copies of a well-conserved 230- to 250-amino-acid cassette called the AAA module that encompasses the Walker A and B motifs. The activities associated with AAA modules are involved in diverse cellular functions.Citation17 Bacterial RuvB, yeast RuvBs (RuvB1 and RuvB2) and human RuvB1 and RuvB2 have been studied under different names. Alternate names of RuvB1/RuvB2 are as RuvBL1/RuvBL2, Pontin/Reptin, TIP49/TIP48, TAP54α/TAP54β, p50/p47, ECP54/ECP51, INO80H/INO80J, TIH1/TIH2, and TIP49A/TIP49B.Citation8,Citation10 RuvB is closely related to clamp-loader subunits. Among the AAA+ class, replication factor-C (RF-C) proteins in eukaryotes and DNA polymerase δ` subunit of E. coli, the clamp-loader subunits play direct roles in loading the clamp proteins for DNA polymerases onto template DNA in an ATP-dependent manner.Citation18
Box 1. Current scenario of prevention and treatment of P. falciparum infection
The current scenario for the prevention and treatment of malaria is becoming difficult due to the emergence of drug resistant P. falciparum parasite. The first case of malarial drug chloroquine resistant parasite was reported in 1957 and consequently chloroquine had become futile for the treatment of malaria. Artemesinin combination therapies are currently being used to treat the uncomplicated P. falciparum malaria in most of the malaria endemic countries (Guidelines for the treatment of malaria, second edition Geneva: WHO, 2011). Recently, it has also been reported that partial artemisinin-resistant P. falciparum malaria has emerged.Citation11 The resistant strains have the potential to spread to different parts of the world and subsequently become a global threat for malaria control and treatment.Citation12 Despite the development of several artemisinin derivative drugs which are being used for the treatment of uncomplicated malariaCitation13 (Guidelines for the treatment of malaria, second edition Geneva: WHO, 2011), the control of malaria in endemic region is limited by drug resistance, relatively high cost and limited availability of newer drugs.Citation14 There are currently no alternative drugs to replace artemisinin derivatives. Considering the future problem and complication of drug resistant parasite, it is necessary to unravel the molecular aspects of pathogenesis of malaria in order to facilitate the development of novel approaches and identification of some novel targets for drug and vaccine candidates. Several efforts have been made to develop vaccine against malaria but their trials failed due to a number of problems while some are still under clinical trials. Like vaccine against malaria, it is equally important to develop next generation antimalarial to treat the drug resistant parasite.
Structural Insight: RuvB/RuvB-like Proteins Exist in Diverse Oligomeric Forms
RuvB can form dimer,Citation19 hexamer,Citation20,Citation21 heptamerCitation21 and dodecamer.Citation22 Such oligomerization is expected to be important for their function together or individually. Diverse oligomeric forms might be required during diverse cellular functions. Analytical ultracentrifugation (AUC) study showed that RuvBL1 and RuvBL2 were mainly monomeric and each monomer co-existed with small proportions of dimer, trimer and hexamer.Citation23 Electron microscopy (EM) studies of RuvBL1/RuvBL2 complex of yeast and human show that RuvBL1 and RuvBL2 form a dodecameric complex consisting of two structurally distinct hexameric rings.Citation22,Citation24 Consistent with this, RuvBL1-RuvBL2 complexes have also been analyzed through AUC and the result showed series of oligomers like monomer, dimers, trimers and single and double hexamers together with smaller forms and the truncation of domain II destabilizes the dodecamer formation.Citation23 The presence of adenine nucleotides induced hexamerization of RuvBL2, but not RuvBL1.Citation23 Gribun et al., proposed a single heterohexameric ring structure for the yeast RuvBL1/RuvBL2 complex.Citation25 The differences between the EM structures suggest that RuvBL1 and RuvBL2 may be capable of forming various complexes.Citation26 Previously, it has been shown that the weak ATPase activity of RuvBL1 and RuvBL2 in vitro increased synergistically when the proteins formed a double-hexameric complex demonstrating that this is the enzymatically active form.Citation22,Citation27 A recent study has confirmed that histidine-tagged yeast RuvB proteins induced the assembly of double hexameric ring of RuvB1/RuvB2 complexes while endogenous RuvB1/RuvB2 complexes are single hexameric rings.Citation28 Bacterial RuvB proteins have a unique β- hairpin protruding from AAA+ ATPase domain which interacts with RuvA protein and play role in branch migration of holliday junctions.Citation29 The hexameric form of RuvB acts as motor protein that drives branch migration by utilizing the energy of ATP hydrolysis.Citation30,Citation31
Biochemical Activities of RuvB Proteins: ATPase and Helicase Activity
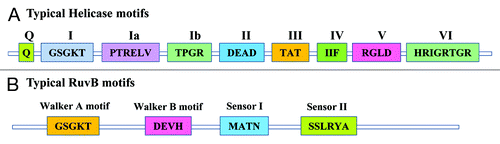
Helicases are ubiquitous enzymes present in prokaryotes, eukaryotes and viruses and are involved in nucleic acid metabolic pathways including DNA replication, repair, recombination and transcription. The universal presence of helicases reflects their fundamental importance in DNA and RNA metabolic processes.Citation32-Citation34 Helicases act on double-stranded nucleic acid substrates,Citation35,Citation36 move directionally along a nucleic acid phosphodiester backbone and separate the two annealed strands by utilizing the energy of ATP hydrolysis.Citation36-Citation38 Helicases are classified on the basis of substrates they utilize and thus helicases can be designated as DNA-DNA, RNA-DNA or RNA-RNA helicases.Citation35,Citation37 Many DNA and RNA helicases possess seven to nine conserved motifs,Citation1 while RuvB is a very unique helicase with respect to its primary structure (). RuvB has no significant overall sequence homologies with them or with helicases with a hexameric ring structure. RuvB contains Walker motifs A and B (), which has been shown to be involved in the binding of ATP and Mg2+. RuvB only displays significant sequence similarity with eukaryotic TIP49a/ RuvBL1 and the related TIP49b/RuvBL2.Citation2,Citation39-Citation41
Figure 2. Comparison of conserved motifs of a typical helicase and RuvB from P. falciparum. (A) Schematics of a typical P. falciparum helicase conserved motifs vs. typical PfRuvB was prepared manually on the basis of different reports 1. P. falciparum helicases contain 7–9 conserved motifs and their conserved amino acid sequences are written in the respective boxes. (B) The basic architecture of PfRuvBs was prepared on the basis of reported conserved motifs in E. coli, yeast and human RuvBs having only Walker A (GSGKT), Walker B (DEAH/DEVH), sensor I and sensor II motifs.
One of the biochemical activities of bacterial RuvBs is their ability to bind and hydrolyze ATP. Yeast RuvB like proteins also bind and hydrolyze ATP as they have conserved Walker A and Walker B motifs. Mutations in Walker A or B motif of either RuvB protein render it inactive.Citation42 It has been shown that recombinant yeast RuvB1/2 complex has enhanced ATPase activity compared with the individual RuvB proteins; furthermore, the ATPase activity of the RuvB1/2 complex is increased in the presence of double-stranded DNA with 5′ or 3′ overhangs. The yeast RuvB1/2 ring undergoes nucleotide-dependent conformational changes as observed by electron microscopy.Citation25 Initially, it was found that both RuvB1 and RuvB2 show ATPase as well as DNA helicase activities.Citation8 It has been established that bacterial RuvB shows 5–3′ direction DNA helicase activity.Citation43 Although it was demonstrated that RuvBs are active as helicases with opposite polarity, some other studies failed to show the same results.Citation40 It has been shown that RuvB1-RuvB2 complex from H. sapiens have ATPase activity but lacked helicase activity.Citation22 RuvB homologs from Mycoplasma pneumoniae and Mycoplasma genitalium showed ATPase activity and divalent cation- and ATP-dependent DNA helicase activity on partially double-stranded DNA substrates.Citation44 Some of the groups have failed to show the ATPase and helicase activities from RuvB protein thus it has been controversial (Box 2). Recombinant human RuvB1 and RuvB2 exhibit lower ATPase activity suggesting that other additional cofactors or substrates may be playing important role in modulating the ATPase activity of the RuvBs.Citation27,Citation45
Box 2. Why biochemical activities of RuvBs are controversial?
The basic architecture of RuvB proteins shows that they do not have all the 7–9 helicase motifs required for the helicase activity. But RuvBs are quite different from helicases having only helicase motif I, II sensor I and sensor II conserved domains and possess ATPase and helicase activity. ATP hydrolyzing property of RuvB protein has become partially controversial as some studies failed to show the ATPase activity. This controversy is due to the experiments showing ATP hydrolysis by recombinant RuvBs which lacked an important negative control, i.e., Walker A or Walker B mutant of these proteins.Citation22,Citation24,Citation25 On the other hand, the failure to detect ATPase activity of recombinant RuvBs could result from inactivation of the protein during purification, /refolding of protein or the requirement of additional factor. Some of the studies show the ATPase activity but failed to establish the DNA helicase activity. The helicase activity of yeast and human RuvBs has become controversial as some groups failed to show the helicase activity and it may be due to the inactivation of the protein during purification/refolding or the requirement of additional cofactors or substrates.Citation27 A recent study has given new insights into the molecular arrangement of RuvBL1 and RuvBL2 and strongly suggested that in vivo activities are regulated by cofactors inducing conformational changes via domain II in order to modulate the enzyme complex into its active state.Citation26
Understanding P. falciparum RuvB Proteins through Bioinformatics Approach
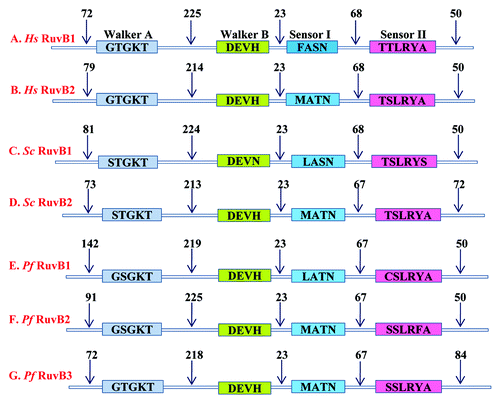
The genome of P. falciparum (www.plasmodb.org) was extensively investigated for the RuvB homologs. The results showed that two homologs of RuvB have been annotated in P. falciparum genome. Recently through genome wide analysis the third homolog of RuvB was identified.Citation1 The PlasmoDB numbers for these are PF08_0100, PF11_0071 and PF13_0330. All of these genes were extensively studied through bioinformatics approaches. The amino acid sequences of all the three PfRuvBs were studied through BlastP analysis. Multiple sequence alignment (MSA) of all the three PfRuvBs, E. coli, S. cerevisiae and H. sapiens RuvB was performed and the results showed that all of these proteins contain motif I and II of a typical helicase. These motifs are also known as Walker motif A and Walker motif B (). Motif I and motif II are required for the ATPase activity of the enzyme.Citation46 On the basis of the results obtained from MSA we have prepared a schematic representation showing conserved motifs of P. falciparum RuvB like protein (). It should be noticed that in all the three PfRuvB proteins, the number of amino acids is highly conserved between various motifs (). PF08_0100 and PF11_0071 contain GSGKT in motif I but PF13_1330 contains GTGKT similar to human, yeast and E. coli. PF08_0100 and PF13_1330 contain DEVH box, similar to human and E. coli RuvB proteins while PF11_0071 contains DEAH sequence in motif II (). The pairwise alignment of PfRuvBs with human RuvB like protein sequence shows 50–43% identity to human homolog.Citation1 PF08_0100, PF11_0071 and PF13_1330 are located on the chromosome number 8, 11 and 13 respectively and encode 520 aa, 475 aa and 483 aa long proteins and their calculated molecular weight ranges from ~53 to 59 kDa.
Figure 3. Conserved motifs of RuvBs from P. falciparum, yeast and human. After multiple sequence alignment of PfRuvBs with yeast and human RuvBs, basic architecture of RuvBs was prepared manually. The name of various motifs with conserved amino acid sequence is written. The numbers denote the N and C-terminal and distances between the conserved motifs.
The phylogenetic analysis of all the three P. falciparum sequences with E. coli, yeast and human was done by using Mac vector with the parameter shown in . The results clearly show that P. falciparum RuvBs are quite diverged from human and E. coli (). On the basis of sequence homology of some conserved motifs and identified role in yeast and other system, it can be expected that P. falciparum RuvBs are AAA+ family of enzymes as scRuvBs and must be playing crucial role in the essential complexes responsible for the survival of parasite.
Figure 4. Phylogenetic analysis of P. falciparum RuvB proteins with other RuvB proteins. The phylogenetic analysis of P. falciparum RuvB like proteins, E. coli, yeast and human RuvBs was carried by using Mac vector with parameters mentioned in the figure.

For structural modeling the sequence of all the P. falciparum RuvBs was submitted to the Swissmodel homology-modeling server (swissmodel.expasy.org/).Citation47 PfRuvB1 primary sequence residues 7 to 513 showed ~42% identity to a human RuvB-like helicase from H. sapiens. PfRuvB2 primary sequence residues 13 to 468 showed ~47% identity to the same human RuvB-like helicase from H. sapiens. Therefore the structural modeling of the PfRuvB1 and PfRuvB2 was done using the known crystal structure of this homolog as the template (PDB number 2C9O at www.rcsb.org). The ribbon diagram of the template is shown in and the predicted structures of PfRuvB1 and PfRuvB2 are shown in and respectively. When the modeled structure of the template and PfRuvB1 or PfRuvB2 were superimposed, it is clear that these structures superimpose partially (, respectively). These results show that the structure of PfRuvB1 and PfRuvB2 are almost similar (). Molecular graphic images were produced using the UCSF Chimera package (www.cgl.ucsf.edu/chimera) from the Resource for Biocomputing, Visualization, and Informatics at the University of California, San Francisco (supported by NIH P41 RR-01081).Citation48
Figure 5. Structure modeling. PfRuvB1 and PfRuvB2 sequences were submitted to Swissmodel server and the structures were obtained. The molecular graphic images were produced using the UCSF Chimera package from the resource for Biocomputing, Visualization, and Informatics (www.cgl.ucsf.edu/chimera) at the University of California, San Francisco (supported by NIH P41 RR-01081). (A) Template for PfRuvB1 and PfRuvB2; (B) PfRuvB1; (C) superimposed image of template and PfRuvB1; d. PfRuvB2; e. superimposed image of template and PfRuvB2.

Functional Insight: RuvB Family of Proteins are Involved in Multiple Biological Processes
It is quite difficult to predict the overall role of RuvB on the basis of sequence homology with other conventional helicases. RuvBs are ubiquitous bacterial proteins with conserved structure and function in distantly related organism.Citation18,Citation49 In prokaryotes the role of some of the interacting partners of RuvB during homologous recombination and replication fork reversal has been established. Several studies performed in yeast and some other system demonstrated that RuvBs are involved in multiple cellular pathways including cell cycle progression and RNA polymerase II-directed transcription.Citation3 RuvBs are also essential components of a wide range of complexes which are involved in the diverse cellular activities. In this section we will describe the involvement of RuvB proteins in diverse cellular processes.
Role of RuvB/RuvBL proteins in cell cycle progression
RuvB containing TIP60 complex, INO80 and SWR1 complexes have been implicated in regulation/activation of cell-cycle checkpoint. Cell cycle checkpoint ensures that the damage is repaired before the progression of the cell cycle to the next stage.Citation50 Activation of cell cycle checkpoints after DNA damages leads to the blockage of cell cycle progression. RuvB1 is required for TIP60s activity and this complex is involved in activation of ATM (Ataxia telangiectasia mutated).Citation51 ATM/ATR (ATM and Rad3-related)-dependent pathways play important role in checkpoint activation which inhibits DNA synthesis quickly.
In higher eukaryotes, RuvBs interact with β-catenin, c-myc, E2F1, and ATF2 which modulate cellular transformation, signaling, apoptosis, DNA damage and response to stress.Citation52-Citation54 The knockdown study of either RuvB1 or RuvB2 in Xenopus has shown embryonic lethality thus these are essential proteins.Citation55 This embryonic lethality has an effect of derepression of the transcription repressor complex c-myc-Miz1.Citation55 Transcription factor Miz1 when complexed with c-myc, inhibits transcription of Miz1 promoter genes. In Xenopus embryo knockdown of either RuvB1 or RuvB2 could be rescued by either inhibition of Miz-1 or overexpression of c-myc. This indicates that RuvB1 and RuvB2 may act as co-repressors of the c-Myc–Miz-1 pathway. The results from investigations of RuvB function in vivo have been more consistent. RuvB1 or RuvB2 knockout from yeast results in non-viability thus these are essential for the survival.Citation2,Citation40,Citation56 Further it has been found that S. cerevisiae RuvB2 protein (Tih2p) is localized to the nucleolus and tightly interacts with Tih1p (RuvB1/TIP49a) which interacts with the TBP and the RNA polymerase II holoenzyme complex. The mutational study of RuvB2 (Tih2p) has confirmed that cells were rapidly arrested in the G1 phase of the cell cycle. Furthermore it has been found that expression of both reptin (RuvBL2) and pontin (RuvBL1) are high during development of mouse.Citation57 The overexpression of either RuvB1 or RuvB2 is toxic in yeast.Citation58 The proteomic analysis of human hepatocellular carcinoma (HCC) has confirmed the overexpression of RuvB2 like (RuvBL2), which is known to interact with β-catenin and cellular v-myc myelocytomatosis viral oncogene homolog (c-myc).Citation59 Recently, silencing experiments in xenografts indicated that RuvBL2 (Reptin) and RuvBL1 (Pontin) are major players in cancer and they may be suitable targets for cancer therapy.Citation60
Involvement of RuvB proteins in mitotic spindle assembly
It has been well identified that RuvBs are involved in regulation of microtubule assembly and organization.Citation61 During mitosis, nucleation of microtubules at the centrosomes and the growth of these microtubules ultimately resulted into the formation of spindle fiber. Various reports have shown the involvement of RuvB1 and RuvB2 in these processes. In human cells, it was found that both RuvB1 and RuvB2 localize to spindles and spindle poles.Citation62,Citation63 Through proteomic approach and a siRNA based screening it has been identified that RuvBs during mitosis function as mitotic regulators.Citation61 The siRNA based reduction of RuvB1 protein levels in HeLa cells resulted in spindle defective phenotypes. RuvBs could indirectly affect spindle formation through interaction with chromatin remodeling complexes.Citation61 The interaction of both RuvB1 and RuvB2 with the g-tubulin ring complex was confirmed by the immunodepletion study of RuvBs in Xenopus extracts. This interaction is implicated in the nucleating spindle formation at centrosomes thus they may be playing direct role in mitotic spindle assembly and organization.Citation61
Involvement of RuvBs in chromatin remodeling
Chromatin-remodeling complexes are extensively involved in facilitating the decompacting of tightly wound DNA for exposure to important factors/complexes such as those involved in transcription, DNA replication, recombination and repair.Citation64 These complexesCitation65 utilize the energy of ATP hydrolysis to mobilize nucleosomes or exchange histones from the DNA or covalently modify histones to change the chromatin state by recruiting proteins that recognize these modifications.Citation64 RuvBs are present in these chromatin-remodeling complexes and play key role in regulation of transcription.
The INO80 complex
The INO80 complex is a large hetero-subunit complex which utilizes the mechanism that mobilizes the nucleosomes.Citation66 The INO80 protein a member of the Swi2/Snf2 superfamily is the catalytically active subunit in INO80 complex and is conserved within eukaryotes.Citation66-Citation69 Both RuvB1 and RuvB2 are present within the INO80 complex in a 6:1 stoichiometry relative to the other subunits of the complex.Citation67 INO80 is a DEAD/H box protein that possesses helicase-type motifs and is an indispensable catalytic component of the INO80 complex.Citation66,Citation67 The analysis of co-immunoprecipitated proteins coupled with mass spectrometry resulted into the identification of RuvB1 and RuvB2.Citation66,Citation70 These results demonstrated that the genes affected by reduction in RuvB2 levels substantially overlapped with those affected in an INO80 knockout strain. In yeast, both INO80 and RuvBs regulate transcription of a similar set of genes, and in vitro assays show that RuvBs are essential for INO80-dependent chromatin-remodeling activity, but not for the binding of INO80 complex to the promoters.Citation67 The purified yeast INO80 complexes were shown to exhibit DNA unwinding activity in an ATP dependent manner, but this activity was abolished when Ino80 was replaced with the Ino80-K737A ATPase-deficient mutant.Citation66 It was suggested that the helicase activity of the RuvBs are functionally coupled to the INO80 ATPase.Citation66 The Ino80 complex purified in the absence of RuvBs was depleted of an actin-like Arp5 subunit that is essential for the chromatin-remodeling activity.Citation67 In vitro association studies indicate that ATP-bound RuvBs promote the assembly of Arp5 into the INO80 complex. Collectively, these results suggest that the primary function of RuvBs is to nucleate the association of Arp5 with the INO80 complex in an ATP-dependent manner. Thus, RuvBs are required for the proper assembly and function of the INO80 complex.
SWR1 or CWR-C chromatin remodeling complex
SWR1 or CWR-C chromatin remodeling complex involves the replacement of histones with histone variants. Swr1 is a Swi2/Snf2- related ATPase and its mammalian homolog is SRCAP. It has been shown that Swr1 complex shows nucleosome stimulated ATPase activity.Citation71 The nucleosome is an octamer of histone proteins and made up of two molecules each of H2A, H2B, H3, and H4 with approximately two turns of DNA (147 bp) wrapped around the octamer.Citation72 Core histones are exchanged with histone variants and these exchanges are with specific DNA-related processes.Citation72,Citation73 In another study it has been confirmed that less H2AX histone variants were recruited to DNA in the absence of SWR-C components.Citation74 Yeast SWR-C or SWR1 complex was studied in detail and found to be composed of 16 subunits including both RuvB1 and RuvB2. Later RuvB1 and RuvB2 were identified in SRCAP (mammalian homolog of SWR-C complex) which is Swi2–Snf2-related CPB activator protein.Citation71,Citation74-Citation76 Swr1 complex shows nucleosome stimulated ATPase activityCitation71 and its ATPase domain plays crucial role in the interaction of Swc2, Swc3, RuvB1, RuvB2, Arp6, and Swc6.Citation77
TIP60 (tat-interactive protein 60) complex
TIP60 (tat-interactive protein 60) complex in human/fly is homologous to NuA4 complex of yeast and is a member of histone acetyltransferase enzymes (HAT) which belong to the MYST (Moz, Ybf2/Sas3, Sas2, and TIP60) family.Citation10 Initially, this complex was purified from yeast and has nucleosomal H4/H2A HAT activity. HATs remodel chromatin structure by acetylating target histones, which results into relaxation of DNA and exposure to proteins located in the vicinity. RuvBs are the essential factors in the mammalian Tip60 complex but not present in the NuA4 complex.Citation27,Citation78 The detailed studies show that both human and yeast complex (TIP60/NuA4) have high degree of conservation.Citation27,Citation79 The constituents of both human TIP60 and yeast NuA4 complexes show the conserved subunits between them and it is clear that RuvB1, RuvB2, YL-l, and homologs of Brd8 are absent in yeast NuA4 complex.Citation10 The catalytic subunit of yeast NuA4 complex and human tip60 complex are Esa1 and TIP60 respectively.Citation27,Citation80
Role of TIP60 complex in transcription and DNA repair
Through various studies it has been established that RuvB1 and RuvB2 act as integral players in the transcriptional processes. The mutational study of RuvB1 and RuvB2 have confirmed that these two proteins are directly or indirectly responsible for the transcription of over 5% of yeast genes, many of which are directly involved in cell cycle regulation.Citation3,Citation9,Citation42 The RuvB1 and RuvB2 containing human TIP60 complex has well established roles in transcription through acetylation of histones at promoters as well as acetylates other cellular proteins involved in transcription such as the androgen receptor (AR). The acetylation of p53 by TIP60 plays crucial role to decide between the cell-cycle arrest and apoptosis.Citation81,Citation82 Initially RuvB1 was identified as a TBP interactor in rat.Citation39 Later RuvB-TBP interaction was confirmed in the yeast, drosophila and human.Citation41,Citation70,Citation83 In addition to TBP, the RuvBs have also been found to interact with various other transcription factors, such as E2F, c-Myc, and β-catenin, all of which play critical role in cell growth, proliferation, and apoptosis.
The TIP60 complex also plays critical roles in DNA damage repair.Citation27 The prerequisite for efficient double-strand break repair is acetylation of histone H2A or variant H2AX which is performed by Tip60 complex.Citation84 In human, RuvB1 is essential for HAT activity of the Tip60 complex.Citation24 The marker for double-stranded breaks is phosphorylated H2AX. Tip60 complex is recruited at this marker and acetylates the histone variant which results in efficient DNA repair.Citation27,Citation78,Citation79,Citation85
Involvement of RuvBs in homologous recombination
Homologous recombination (HR) is an essential process during which gene rearrangement in meiosis and double-strand break repair of stalled replication forks takes place.Citation86-Citation88 The AAA+ domain of yeast RuvB1 and RuvB2 are weakly homologous to the bacterial RuvB helicase. Both bacterial and yeast RuvB helicases are involved in the process of HR.Citation89,Citation90 By the process of HR, organism attains genomic integrity and genetic diversity.Citation91 During HR in prokaryotes RuvA, RuvB, and RuvC proteins play key role in branch migration and resolution of holliday junction.Citation30,Citation31,Citation92-Citation95 Structural and mutational study of RuvB suggested that RuvB is a multifunctional protein which interacts with RuvA, ATP, DNA, and Mg2+ and all these interactions are required for RuvAB dependent branch migration of the holliday junction. The RuvB hexamer is a motor and energy required for the branch migration is provided by the ATPase activity of RuvB and this activity is synergistically enhanced by RuvA and DNA.Citation96 Takashi et al., have established that the point mutation in Walker A motif greatly affects the diverse activities of RuvB such as ATP hydrolysis, ATP binding, DNA binding, hexamer formation and promotion of branch migration.Citation97
Involvement of RuvBs protein in Replication Fork Reversal (RFR)
Often in prokaryotes, DNA replication can be impaired by obstacle or inactivation of replication proteins resulting into replication arrest. The consequences of replication arrest are highly dramatic and now recognized as major source of genomic instability in all organisms.Citation98-Citation102 There are reports from replication mutants which show that a specific reaction takes place prior to replication restart called RFR.Citation4,Citation103 The primary step of RFR is the annealing of the leading- and lagging-strand ends which results in the formation of a holliday junction adjacent to DNA double-strand end, both of which are processed by recombination enzymes. RFR is catalyzed by RuvAB complex in several replication mutants originally characterized for its role in the last steps of HR, branch migration and resolution of holliday junction.Citation104 RFR has been well studied in E. coli.Citation18,Citation90 RecBCD is a recombination complex specific for the repair of dsDNA breaks which act on ds DNA ends. This complex gradually unwinds the duplex and degrades up to a specific site named Chi. RecBCD at Chi shifts to a recombinase and loads the RecA recombination enzyme.Citation105 Holliday junctions made by fork reversal are branch-migrated by the RuvAB complex and later RuvABC complex act as resolvase and resolve the holliday junction.Citation95
Involvement of RuvBs in snoRNPs biogenesis and assembly
The biogenesis and assembly of the small nucleolar ribonucleoproteins complex (snoRNPs) involves the transcription of the snoRNA with the assembly and restructuring of the core proteins interacting with the snoRNAs. After mature snoRNPs are formed, they participate in the modification and processing of pre-rRNA in the nucleolus.Citation8 RuvBs have also been studied in terms of the biogenesis and assembly of the snoRNP in yeast and human in detail. RuvBs association with snoRNPs has been well established. snoRNPs are crucial for the cleavage and modification of small nuclear RNA (sRNA), rRNA (rRNA), and tRNAs (tRNA).Citation106 The depletion study of yeast RuvB2 by use of a galactose-regulated allele of RuvB2 decreased the levels of snoRNAs.Citation107 In human cell lines, siRNAs mediated depletion of endogenous RuvB proteins led to the decreased levels of snoRNAs.Citation7 RuvB1 and RuvB2 interact and form a tight complex with Pih1 (also called Nop17, a Nop5/Nop58 interacting protein). In yeast, Pih1 deletion resulted into accumulation of the 35S pre-rRNA.Citation108,Citation109 Thus this result suggests the role of Pih1 in pre rRNA processing and involvement in snoRNP biogenesis. Additionally it has been shown that RuvB2-GFP localizes to the nucleolus. The above experimental results indicate that RuvBs must be playing important role in the assembly and localization of snoRNPs and stability of snoRNAs.
snoRNPs are ribonucleolar proteins complexes and have been classified into three families like box C/D snoRNP, box H/ACA snoRNPs and the MRP snoRNPs. So far association of snoRNPs of the box C/D with RuvBs has been studied in detail.Citation7,Citation107,Citation109-Citation112 In vitro interaction between RuvBs with immobilized U14 and U15 RNAs (examples of box C/D snoRNAs) have been found.Citation110 Nop1 is a core snoRNP protein that associates with snoRNAs of box C/D family. In a recent study it has been demonstrated that RuvB1 (TIP48) and RuvB2 (TIP49) efficiently bridge interactions between the core box C/D proteins NOP56 or NOP58 and 15.5K.Citation113 Gar1 is a core snoRNP protein that associates with snoRNAs of H/ACA family. Both nop1 and gar1 are localized to the nucleolus, the site of rRNA gene transcription and processing. It has been shown that the depletion of RuvB2 results into the dispersion of both Nop1 and Gar1 from nucleolus to the nucleoplasm. These results suggest that RuvB2 is required for localization of nop1 and gar1 to the nucleolus and is probably involved in the assembly of the snoRNP.Citation10
Involvement of RuvBs in telomerase core complex assembly
Another important role of RuvB protein has been found in the assembly of the telomerase core complex. Telomerase is an enzyme that adds DNA sequence repeats to the 3′ end of DNA strands in the telomere regions/ends of the chromosomes. The repeated nucleotide region is called telomeres which contains non-coding DNA and prevents constant loss of important DNA from chromosome ends thus stabilizing the chromosome. The telomerase core complex is made up of ribonucleoprotein particle, consisting of three essential components: TERT (the telomerase reverse transcriptase), dyskerin (a core box H/ACA RNA-binding protein) and TERC (Telomerase RNA component).Citation114,Citation115 Pontin (RuvB1) and Reptin (RuvB2) have been identified as telomerase components which are essential for holoenzyme assembly.Citation116 Both RuvB1 and RuvB2 individually interact with TERT and dyskerin in a TERC-independent manner.Citation116 The depletion study of RuvBs showed reduced dyskerin levels which are required to stabilize TERC. Additionally it has been found that the loss of RuvBs resulted into decreased telomerase activity by 80–90% due to a decrease in TERC RNA.Citation10 During S phase of the cell cycle, the association of RuvBs with the TERT complex increases suggesting their role in maintaining telomerase activity when the function of telomerase is most critical.Citation10 The identification of TERT-RuvB1-RuvB2 complex as a stable complex suggests that the RuvBs possibly bridge TERT to the dyskerin-TERC particle which further leads to the formation of the TERT-dyskerin-TERC core telomerase complex.Citation116 It has been shown that TERT/RuvB1/RuvB2 complex has decreased telomerase activity which indicates that RuvB proteins are present in a pretelomerase complex that promotes the formation of an active telomerase complex and dissociation of RuvBs occurs. Hence it has been proposed that RuvBs act as scaffolding protein required for the formation of active complex or functions as remodeling proteins for the maturation of the telomerase core complex.
Predicted PfRuvB Containing Complexes and Their Role
RuvBs have been found to interact with several proteins associated with diverse cellular activities in the E. coli, yeast, drosophila and human. Initially in order to find the interacting partners of P. falciparum RuvBs we have performed similarity search with the known proteins of E. coli, yeast, drosophila and human etc. As the RuvBs have been found to play critical role in chromatin remodeling, the P. falciparum genome was investigated for Chromatin Remodeling Protein (CRP) and the result showed that several putative CRPs are present. Further investigation for RuvB interacting swi-snf related proteins of chromatin remodeling complexes like INO80 complex and SWR1 complex (yeast)/SRCAP (SWI/SNF-related CBP activator protein in mammal) was also done. In order to find INO80 like protein in P. falciparum, INO80 protein sequence from different organism was retrieved and BlastP analysis with PlasmoDB shows that at least three proteins with high blast score are present and their PlasmoDB numbers are PF08_0048, PFF1188w and PFF0560. PlasmoDB does not have any annotated SWR1/SWR-C (present in yeast) protein yet. But there is one Swi-Snf related CBP activator gene (PF08_0048) present which seems homologous to SRCAP (homologous to swr1 complex in mammal). MSA of this protein with human SWR1 and yeast SRCAP protein showed that this P. falciparum protein has few conserved domains as in human and yeast. Thus the presence of INO80 and SRCAP like proteins in P. falciparum, suggests that both these complexes possibly exist in the P. falciparum.
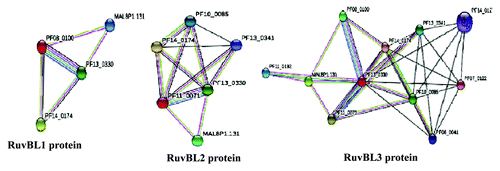
TIP60/NuA4 is basically histone acetyltransferase and one gene with high blast score (PF11_0192) is present in P. falciparum genome. Besides the presence of these key regulators of CRC, there are several other P. falciparum specific chromatin remodeling proteins present in the PlasmoDB which may be part of the unexplored CRC of P. falciparum in which RuvBs probably play essential role. After searching the whole genome of P. falciparum we found some other proteins which either interact with PfRuvBs or are similar to the known proteins of E. coli, yeast and human. The PlasmoDB number of these proteins are PfL2215w (actin1 like protein), arp4 homolog (Pf14_0218), arp6 (Pf07_0077) and arp5 like protein (PfA0190c), Pf14_0068 (putative Nop1/fibrilarian protein/crucial box C/D protein), Pf13_0051 (snoRNP protein, homolog of Gar1- belongs to box H/ACA), Pf13_0080 (telomerase reverse transcriptase/TERT). Well-known RuvB-interacting protein β-catenin homolog is also present in the P. falciparum. Other RuvB interacting proteins like TBP and E2F like proteins are also present in the P. falciparum genome. Further to refine the results we have used bioinformatics approach to identify the interacting partners and the results show that PfRuvB1 interacts with PfRuvB3, a snoRNP (Pf14_0174) and Gas41 homolog (MAL8P1.131) (). PfRuvB2 interacts with PfRuvB3, Nop5 homolog (Pf10_0085), a snoRNP (Pf14_0174) and also with DNA directed RNA polymerase II (Pf13_0341) (). PfRuvB3 seems to be interacting with more proteins than PfRuvB1 or PfRuvB2 and the results showed that it interacts with at least nine proteins (). The PlasmoDB numbers of these proteins are PF14_0174 (a snoRNP), PfRuvB2, MALP1.131 (Gas41 homolog), PF10_0085 (Nop5/Nop58 homolog), PF13_0341 (DNA directed RNA polymerase-II), PF11_0192 (Pf specific histone acetyl transferase), PF08_0041 (ribosome biogenesis protein nep1 homolog, putative), Pf14_0127 (N-myristoyl transferase) and Pf07_0122 (putative BRIX domain protein). The interactomes were developed by using STRING 9.0 ().
Figure 6. Prediction of P. falciparum RuvB proteins interacting proteins. P. falciparum RuvB proteins amino acid sequences were analyzed with STRING 9.0 at Expasy (expasy.org). The results showed the presence of at least three proteins interacting with PfRuvB1, four proteins interacting with PfRuvB2 and nine proteins interacting with PfRuvB3. The PlasmoDB numbers of all are shown in the model. This analysis showed the interaction of PfRuvB3 with PfRuvB1 and PfRuvB2 proteins. Nop5 also interacts with both PfRuvB2 and PfRuvB3 proteins. Gas41 homolog seems to be interacting with all the three PfRuvB proteins.
On the basis of the presence of crucial RuvB interacting homologs in the P. falciparum genome, it can be expected that P. falciparum RuvBs are ATPase of AAA+ family of enzymes and are most likely involved in the diverse cellular activities including chromatin remodeling and cell cycle progression. Thus it will be interesting to explore the detailed components of PfRuvB complexes which may further lead to the identification of some novel P. falciparum specific RuvB interacting proteins.
Concluding Remarks
RuvBs are ubiquitous proteins belonging to AAA+ class of enzymes present in bacteria, yeast, drosophila, human, Plasmodium etc and are essential for the survival of yeast and drosophila. Biochemical activities especially helicase activity has been controversial as some of the groups have failed to establish the helicase activity from recombinant proteins. This failure may be due to the inactivation of the protein during purification/refolding or the requirement of additional cofactors or substrates.Citation10,Citation27,Citation117 To overcome from this controversy it has been necessary to establish their biochemical activity from purified endogenous protein. Further biological role of PfRuvB complex has to be established in order to understand their importance in the parasite life cycle. MSA of PfRuvBs confirmed the presence of typical RuvB conserved domains thus there is a high probability that these proteins contain the ATPase activity and possibly helicase activity. The phylogenetic analysis clearly shows that PfRuvBs are quite diverged from human and E. coli and are closer to yeast RuvBs which are essential for the survival of yeast. It is also expected that P. falciparum RuvBs must be playing crucial role in the multiprotein complexes like CRC, snoRNPs biogenesis complexes and multiprotein complexes essential for cell cycle progression etc. Considering all the above facts and expectations it is interesting to characterize P. falciparum RuvBs in detail which can serve as a very good drug target (Box 3) if they show specific and essential role in the survival of the parasite.
Box 3. Why PfRuvB proteins are expected as a potential drug target?
In order to expect/establish a gene/protein as a potential drug target, the gene/protein should be parasite specific and essential for the survival. A recent report on genome wide analysis of P. falciparum helicases suggested that parasite specific helicases can be good antimalarial drug targets.Citation1 The knockout study of scRuvB proteins have confirmed its requirement for the survival of yeast and similar results were also seen in the survival of drosophila. Furthermore, scRuvB have been found essential for the cell cycle progression. The knockdown study of either RuvB1 or RuvB2 in Xenopus has shown embryonic lethality thus these are essential proteins.Citation55 In vivo silencing study of Reptin (RuvBL2) blocks the progression of human hepatocellular carcinoma and it has been suggested as a suitable drug target for cancer.Citation16 The analysis of amino acid sequence of PfRuvBs and the phylogenetic analysis confirmed the presence of conserved domains of a typical RuvB which is phylogenetically closer to the yeast RuvBs as compared with the human and E. coli. Thus on the basis of above studies and finding, PfRuvBs can be highly expected as essential for the parasite survival and may be suitable drug target to develop new antimalarials.
Acknowledgments
We apologize to all authors whose contributions could not be acknowledged due to space limitations. The work in RT’s laboratory is partially supported by Department of Biotechnology, Department of Science and Technology and Defense Research and Development Organization grants. Infrastructural support from the Department of Biotechnology, Government of India, is gratefully acknowledged.
Related Research Data
References
- Tuteja R. Genome wide identification of Plasmodium falciparum helicases: a comparison with human host. Cell Cycle 2010; 9:104 - 20; http://dx.doi.org/10.4161/cc.9.1.10241; PMID: 20016272
- Qiu XB, Lin YL, Thome KC, Pian P, Schlegel BP, Weremowicz S, et al. An eukaryotic RuvB-like protein (RUVBL1) essential for growth. J Biol Chem 1998; 273:27786 - 93; http://dx.doi.org/10.1074/jbc.273.43.27786; PMID: 9774387
- Lim CR, Kimata Y, Ohdate H, Kokubo T, Kikuchi N, Horigome T, et al. The Saccharomyces cerevisiae RuvB-like protein, Tih2p, is required for cell cycle progression and RNA polymerase II-directed transcription. J Biol Chem 2000; 275:22409 - 17; http://dx.doi.org/10.1074/jbc.M001031200; PMID: 10787406
- Seigneur M, Bidnenko V, Ehrlich SD, Michel B. RuvAB acts at arrested replication forks. Cell 1998; 95:419 - 30; http://dx.doi.org/10.1016/S0092-8674(00)81772-9; PMID: 9814711
- Izumi N, Yamashita A, Iwamatsu A, Kurata R, Nakamura H, Saari B, et al. AAA+ proteins RUVBL1 and RUVBL2 coordinate PIKK activity and function in nonsense-mediated mRNA decay. Sci Signal 2010; 3:ra27; http://dx.doi.org/10.1126/scisignal.2000468; PMID: 20371770
- Xie X, Chen Y, Xue P, Fan Y, Deng Y, Peng G, et al. RUVBL2, a novel AS160-binding protein, regulates insulin-stimulated GLUT4 translocation. Cell Res 2009; 19:1090 - 7; http://dx.doi.org/10.1038/cr.2009.68; PMID: 19532121
- Watkins NJ, Lemm I, Ingelfinger D, Schneider C, Hossbach M, Urlaub H, et al. Assembly and maturation of the U3 snoRNP in the nucleoplasm in a large dynamic multiprotein complex. Mol Cell 2004; 16:789 - 98; http://dx.doi.org/10.1016/j.molcel.2004.11.012; PMID: 15574333
- Huen J, Kakihara Y, Ugwu F, Cheung KL, Ortega J, Houry WA. Rvb1-Rvb2: essential ATP-dependent helicases for critical complexes. Biochem Cell Biol 2010; 88:29 - 40; http://dx.doi.org/10.1139/O09-122; PMID: 20130677
- Wood MA, McMahon SB, Cole MD. An ATPase/helicase complex is an essential cofactor for oncogenic transformation by c-Myc. Mol Cell 2000; 5:321 - 30; http://dx.doi.org/10.1016/S1097-2765(00)80427-X; PMID: 10882073
- Jha S, Dutta A. RVB1/RVB2: running rings around molecular biology. Mol Cell 2009; 34:521 - 33; http://dx.doi.org/10.1016/j.molcel.2009.05.016; PMID: 19524533
- Noedl H, Socheat D, Satimai W. Artemisinin-resistant malaria in Asia. N Engl J Med 2009; 361:540 - 1; http://dx.doi.org/10.1056/NEJMc0900231; PMID: 19641219
- Dondorp AM, Yeung S, White L, Nguon C, Day NP, Socheat D, et al. Artemisinin resistance: current status and scenarios for containment. Nat Rev Microbiol 2010; 8:272 - 80; http://dx.doi.org/10.1038/nrmicro2385; PMID: 20208550
- Baird JK. Effectiveness of antimalarial drugs. N Engl J Med 2005; 352:1565 - 77; http://dx.doi.org/10.1056/NEJMra043207; PMID: 15829537
- Rosenthal PJ. Artesunate for the treatment of severe falciparum malaria. N Engl J Med 2008; 358:1829 - 36; http://dx.doi.org/10.1056/NEJMct0709050; PMID: 18434652
- Li W, Zeng J, Li Q, Zhao L, Liu T, Björkholm M, et al. Reptin is required for the transcription of telomerase reverse transcriptase and over-expressed in gastric cancer. Mol Cancer 2010; 9:132; http://dx.doi.org/10.1186/1476-4598-9-132; PMID: 20509972
- Ménard L, Taras D, Grigoletto A, Haurie V, Nicou A, Dugot-Senant N, et al. In vivo silencing of Reptin blocks the progression of human hepatocellular carcinoma in xenografts and is associated with replicative senescence. J Hepatol 2010; 52:681 - 9; http://dx.doi.org/10.1016/j.jhep.2009.12.029; PMID: 20346530
- Neuwald AF, Aravind L, Spouge JL, Koonin EV. AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res 1999; 9:27 - 43; PMID: 9927482
- Iwasaki H, Han YW, Okamoto T, Ohnishi T, Yoshikawa M, Yamada K, et al. Mutational analysis of the functional motifs of RuvB, an AAA+ class helicase and motor protein for holliday junction branch migration. Mol Microbiol 2000; 36:528 - 38; http://dx.doi.org/10.1046/j.1365-2958.2000.01842.x; PMID: 10844644
- Shiba T, Iwasaki H, Nakata A, Shinagawa H. Escherichia coli RuvA and RuvB proteins involved in recombination repair: physical properties and interactions with DNA. Mol Gen Genet 1993; 237:395 - 9; PMID: 8483454
- Mitchell AH, West SC. Hexameric rings of Escherichia coli RuvB protein. Cooperative assembly, processivity and ATPase activity. J Mol Biol 1994; 243:208 - 15; http://dx.doi.org/10.1006/jmbi.1994.1648; PMID: 7932751
- Miyata T, Yamada K, Iwasaki H, Shinagawa H, Morikawa K, Mayanagi K. Two different oligomeric states of the RuvB branch migration motor protein as revealed by electron microscopy. J Struct Biol 2000; 131:83 - 9; http://dx.doi.org/10.1006/jsbi.2000.4290; PMID: 11042078
- Puri T, Wendler P, Sigala B, Saibil H, Tsaneva IR. Dodecameric structure and ATPase activity of the human TIP48/TIP49 complex. J Mol Biol 2007; 366:179 - 92; http://dx.doi.org/10.1016/j.jmb.2006.11.030; PMID: 17157868
- Niewiarowski A, Bradley AS, Gor J, McKay AR, Perkins SJ, Tsaneva IR. Oligomeric assembly and interactions within the human RuvB-like RuvBL1 and RuvBL2 complexes. Biochem J 2010; 429:113 - 25; http://dx.doi.org/10.1042/BJ20100489; PMID: 20412048
- Torreira E, Jha S, López-Blanco JR, Arias-Palomo E, Chacón P, Cañas C, et al. Architecture of the pontin/reptin complex, essential in the assembly of several macromolecular complexes. Structure 2008; 16:1511 - 20; http://dx.doi.org/10.1016/j.str.2008.08.009; PMID: 18940606
- Gribun A, Cheung KL, Huen J, Ortega J, Houry WA. Yeast Rvb1 and Rvb2 are ATP-dependent DNA helicases that form a heterohexameric complex. J Mol Biol 2008; 376:1320 - 33; http://dx.doi.org/10.1016/j.jmb.2007.12.049; PMID: 18234224
- Gorynia S, Bandeiras TM, Pinho FG, McVey CE, Vonrhein C, Round A, et al. Structural and functional insights into a dodecameric molecular machine—the RuvBL1/RuvBL2 complex. J Struct Biol 2011; 176:279 - 91; http://dx.doi.org/10.1016/j.jsb.2011.09.001; PMID: 21933716
- Ikura T, Ogryzko VV, Grigoriev M, Groisman R, Wang J, Horikoshi M, et al. Involvement of the TIP60 histone acetylase complex in DNA repair and apoptosis. Cell 2000; 102:463 - 73; http://dx.doi.org/10.1016/S0092-8674(00)00051-9; PMID: 10966108
- Cheung KL, Huen J, Houry WA, Ortega J. Comparison of the multiple oligomeric structures observed for the Rvb1 and Rvb2 proteins. Biochem Cell Biol 2010; 88:77 - 88; http://dx.doi.org/10.1139/O09-159; PMID: 20130681
- Han YW, Iwasaki H, Miyata T, Mayanagi K, Yamada K, Morikawa K, et al. A unique beta-hairpin protruding from AAA+ ATPase domain of RuvB motor protein is involved in the interaction with RuvA DNA recognition protein for branch migration of Holliday junctions. J Biol Chem 2001; 276:35024 - 8; http://dx.doi.org/10.1074/jbc.M103611200; PMID: 11427534
- Iwasaki H, Takahagi M, Nakata A, Shinagawa H. Escherichia coli RuvA and RuvB proteins specifically interact with Holliday junctions and promote branch migration. Genes Dev 1992; 6:2214 - 20; http://dx.doi.org/10.1101/gad.6.11.2214; PMID: 1427081
- Tsaneva IR, Müller B, West SC. ATP-dependent branch migration of Holliday junctions promoted by the RuvA and RuvB proteins of E. coli. Cell 1992; 69:1171 - 80; http://dx.doi.org/10.1016/0092-8674(92)90638-S; PMID: 1617728
- Matson SW, Bean DW, George JW. DNA helicases: enzymes with essential roles in all aspects of DNA metabolism. Bioessays 1994; 16:13 - 22; http://dx.doi.org/10.1002/bies.950160103; PMID: 8141804
- Hall MC, Matson SW. Helicase motifs: the engine that powers DNA unwinding. Mol Microbiol 1999; 34:867 - 77; http://dx.doi.org/10.1046/j.1365-2958.1999.01659.x; PMID: 10594814
- Vindigni A. Biochemical, biophysical, and proteomic approaches to study DNA helicases. Mol Biosyst 2007; 3:266 - 74; PMID: 17372655
- Tuteja R. Helicases - feasible antimalarial drug target for Plasmodium falciparum. FEBS J 2007; 274:4699 - 704; http://dx.doi.org/10.1111/j.1742-4658.2007.06000.x; PMID: 17824956
- Tuteja N, Tuteja R. Unraveling DNA helicases. Motif, structure, mechanism and function. Eur J Biochem 2004; 271:1849 - 63; http://dx.doi.org/10.1111/j.1432-1033.2004.04094.x; PMID: 15128295
- Tuteja N, Tuteja R. Prokaryotic and eukaryotic DNA helicases. Essential molecular motor proteins for cellular machinery. Eur J Biochem 2004; 271:1835 - 48; http://dx.doi.org/10.1111/j.1432-1033.2004.04093.x; PMID: 15128294
- Linder P, Lasko P. Bent out of shape: RNA unwinding by the DEAD-box helicase Vasa. Cell 2006; 125:219 - 21; http://dx.doi.org/10.1016/j.cell.2006.03.030; PMID: 16630807
- Kanemaki M, Makino Y, Yoshida T, Kishimoto T, Koga A, Yamamoto K, et al. Molecular cloning of a rat 49-kDa TBP-interacting protein (TIP49) that is highly homologous to the bacterial RuvB. Biochem Biophys Res Commun 1997; 235:64 - 8; http://dx.doi.org/10.1006/bbrc.1997.6729; PMID: 9196036
- Kanemaki M, Kurokawa Y, Matsu-ura T, Makino Y, Masani A, Okazaki K, et al. TIP49b, a new RuvB-like DNA helicase, is included in a complex together with another RuvB-like DNA helicase, TIP49a. J Biol Chem 1999; 274:22437 - 44; http://dx.doi.org/10.1074/jbc.274.32.22437; PMID: 10428817
- Makino Y, Mimori T, Koike C, Kanemaki M, Kurokawa Y, Inoue S, et al. TIP49, homologous to the bacterial DNA helicase RuvB, acts as an autoantigen in human. Biochem Biophys Res Commun 1998; 245:819 - 23; http://dx.doi.org/10.1006/bbrc.1998.8504; PMID: 9588198
- Jónsson ZO, Dhar SK, Narlikar GJ, Auty R, Wagle N, Pellman D, et al. Rvb1p and Rvb2p are essential components of a chromatin remodeling complex that regulates transcription of over 5% of yeast genes. J Biol Chem 2001; 276:16279 - 88; http://dx.doi.org/10.1074/jbc.M011523200; PMID: 11278922
- Tsaneva IR, Müller B, West SC. RuvA and RuvB proteins of Escherichia coli exhibit DNA helicase activity in vitro. Proc Natl Acad Sci U S A 1993; 90:1315 - 9; http://dx.doi.org/10.1073/pnas.90.4.1315; PMID: 8433990
- Estevão S, Sluijter M, Hartwig NG, van Rossum AM, Vink C. Functional characterization of the RuvB homologs from Mycoplasma pneumoniae and Mycoplasma genitalium. J Bacteriol 2011; 193:6425 - 35; http://dx.doi.org/10.1128/JB.06003-11; PMID: 21949077
- Fuchs M, Gerber J, Drapkin R, Sif S, Ikura T, Ogryzko V, et al. The p400 complex is an essential E1A transformation target. Cell 2001; 106:297 - 307; http://dx.doi.org/10.1016/S0092-8674(01)00450-0; PMID: 11509179
- Tuteja R, Pradhan A. Unraveling the ‘DEAD-box’ helicases of Plasmodium falciparum. Gene 2006; 376:1 - 12; http://dx.doi.org/10.1016/j.gene.2006.03.007; PMID: 16713133
- Arnold K, Bordoli L, Kopp J, Schwede T. The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. Bioinformatics 2006; 22:195 - 201; http://dx.doi.org/10.1093/bioinformatics/bti770; PMID: 16301204
- Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, et al. UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 2004; 25:1605 - 12; http://dx.doi.org/10.1002/jcc.20084; PMID: 15264254
- Rocha EP, Cornet E, Michel B. Comparative and evolutionary analysis of the bacterial homologous recombination systems. PLoS Genet 2005; 1:e15; http://dx.doi.org/10.1371/journal.pgen.0010015; PMID: 16132081
- Sancar A, Lindsey-Boltz LA, Unsal-Kaçmaz K, Linn S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu Rev Biochem 2004; 73:39 - 85; http://dx.doi.org/10.1146/annurev.biochem.73.011303.073723; PMID: 15189136
- Sun Y, Jiang X, Chen S, Fernandes N, Price BD. A role for the Tip60 histone acetyltransferase in the acetylation and activation of ATM. Proc Natl Acad Sci U S A 2005; 102:13182 - 7; http://dx.doi.org/10.1073/pnas.0504211102; PMID: 16141325
- Bauer A, Chauvet S, Huber O, Usseglio F, Rothbächer U, Aragnol D, et al. Pontin52 and reptin52 function as antagonistic regulators of beta-catenin signalling activity. EMBO J 2000; 19:6121 - 30; http://dx.doi.org/10.1093/emboj/19.22.6121; PMID: 11080158
- Cho SG, Bhoumik A, Broday L, Ivanov V, Rosenstein B, Ronai Z. TIP49b, a regulator of activating transcription factor 2 response to stress and DNA damage. Mol Cell Biol 2001; 21:8398 - 413; http://dx.doi.org/10.1128/MCB.21.24.8398-8413.2001; PMID: 11713276
- Dugan KA, Wood MA, Cole MD. TIP49, but not TRRAP, modulates c-Myc and E2F1 dependent apoptosis. Oncogene 2002; 21:5835 - 43; http://dx.doi.org/10.1038/sj.onc.1205763; PMID: 12185582
- Etard C, Gradl D, Kunz M, Eilers M, Wedlich D. Pontin and Reptin regulate cell proliferation in early Xenopus embryos in collaboration with c-Myc and Miz-1. Mech Dev 2005; 122:545 - 56; http://dx.doi.org/10.1016/j.mod.2004.11.010; PMID: 15804567
- Radovic S, Rapisarda VA, Tosato V, Bruschi CV. Functional and comparative characterization of Saccharomyces cerevisiae RVB1 and RVB2 genes with bacterial Ruv homologues. FEMS Yeast Res 2007; 7:527 - 39; http://dx.doi.org/10.1111/j.1567-1364.2006.00205.x; PMID: 17302941
- Chauvet S, Usseglio F, Aragnol D, Pradel J. Analysis of paralogous pontin and reptin gene expression during mouse development. Dev Genes Evol 2005; 215:575 - 9; http://dx.doi.org/10.1007/s00427-005-0011-1; PMID: 16003523
- Sopko R, Huang D, Preston N, Chua G, Papp B, Kafadar K, et al. Mapping pathways and phenotypes by systematic gene overexpression. Mol Cell 2006; 21:319 - 30; http://dx.doi.org/10.1016/j.molcel.2005.12.011; PMID: 16455487
- Rousseau B, Ménard L, Haurie V, Taras D, Blanc JF, Moreau-Gaudry F, et al. Overexpression and role of the ATPase and putative DNA helicase RuvB-like 2 in human hepatocellular carcinoma. Hepatology 2007; 46:1108 - 18; http://dx.doi.org/10.1002/hep.21770; PMID: 17657734
- Grigoletto A, Lestienne P, Rosenbaum J. The multifaceted proteins Reptin and Pontin as major players in cancer. Biochim Biophys Acta 2011; 1815:147 - 57; PMID: 21111787
- Ducat D, Kawaguchi S, Liu H, Yates JR 3rd, Zheng Y. Regulation of microtubule assembly and organization in mitosis by the AAA+ ATPase Pontin. Mol Biol Cell 2008; 19:3097 - 110; http://dx.doi.org/10.1091/mbc.E07-11-1202; PMID: 18463163
- Gartner W, Rossbacher J, Zierhut B, Daneva T, Base W, Weissel M, et al. The ATP-dependent helicase RUVBL1/TIP49a associates with tubulin during mitosis. Cell Motil Cytoskeleton 2003; 56:79 - 93; http://dx.doi.org/10.1002/cm.10136; PMID: 14506706
- Sigala B, Edwards M, Puri T, Tsaneva IR. Relocalization of human chromatin remodeling cofactor TIP48 in mitosis. Exp Cell Res 2005; 310:357 - 69; http://dx.doi.org/10.1016/j.yexcr.2005.07.030; PMID: 16157330
- Ruthenburg AJ, Li H, Patel DJ, Allis CD. Multivalent engagement of chromatin modifications by linked binding modules. Nat Rev Mol Cell Biol 2007; 8:983 - 94; http://dx.doi.org/10.1038/nrm2298; PMID: 18037899
- Narlikar GJ, Fan HY, Kingston RE. Cooperation between complexes that regulate chromatin structure and transcription. Cell 2002; 108:475 - 87; http://dx.doi.org/10.1016/S0092-8674(02)00654-2; PMID: 11909519
- Shen X, Mizuguchi G, Hamiche A, Wu C. A chromatin remodelling complex involved in transcription and DNA processing. Nature 2000; 406:541 - 4; http://dx.doi.org/10.1038/35020123; PMID: 10952318
- Jónsson ZO, Jha S, Wohlschlegel JA, Dutta A. Rvb1p/Rvb2p recruit Arp5p and assemble a functional Ino80 chromatin remodeling complex. Mol Cell 2004; 16:465 - 77; http://dx.doi.org/10.1016/j.molcel.2004.09.033; PMID: 15525518
- Jin J, Cai Y, Yao T, Gottschalk AJ, Florens L, Swanson SK, et al. A mammalian chromatin remodeling complex with similarities to the yeast INO80 complex. J Biol Chem 2005; 280:41207 - 12; http://dx.doi.org/10.1074/jbc.M509128200; PMID: 16230350
- Bao Y, Shen X. INO80 subfamily of chromatin remodeling complexes. Mutat Res 2007; 618:18 - 29; http://dx.doi.org/10.1016/j.mrfmmm.2006.10.006; PMID: 17316710
- Ohdate H, Lim CR, Kokubo T, Matsubara K, Kimata Y, Kohno K. Impairment of the DNA binding activity of the TATA-binding protein renders the transcriptional function of Rvb2p/Tih2p, the yeast RuvB-like protein, essential for cell growth. J Biol Chem 2003; 278:14647 - 56; http://dx.doi.org/10.1074/jbc.M213220200; PMID: 12576485
- Mizuguchi G, Shen X, Landry J, Wu WH, Sen S, Wu C. ATP-driven exchange of histone H2AZ variant catalyzed by SWR1 chromatin remodeling complex. Science 2004; 303:343 - 8; http://dx.doi.org/10.1126/science.1090701; PMID: 14645854
- Sarma K, Reinberg D. Histone variants meet their match. Nat Rev Mol Cell Biol 2005; 6:139 - 49; http://dx.doi.org/10.1038/nrm1567; PMID: 15688000
- Malik HS, Henikoff S. Phylogenomics of the nucleosome. Nat Struct Biol 2003; 10:882 - 91; http://dx.doi.org/10.1038/nsb996; PMID: 14583738
- Krogan NJ, Keogh MC, Datta N, Sawa C, Ryan OW, Ding H, et al. A Snf2 family ATPase complex required for recruitment of the histone H2A variant Htz1. Mol Cell 2003; 12:1565 - 76; http://dx.doi.org/10.1016/S1097-2765(03)00497-0; PMID: 14690608
- Kobor MS, Venkatasubrahmanyam S, Meneghini MD, Gin JW, Jennings JL, Link AJ, et al. A protein complex containing the conserved Swi2/Snf2-related ATPase Swr1p deposits histone variant H2A.Z into euchromatin. PLoS Biol 2004; 2:E131; http://dx.doi.org/10.1371/journal.pbio.0020131; PMID: 15045029
- Cai Y, Jin J, Florens L, Swanson SK, Kusch T, Li B, et al. The mammalian YL1 protein is a shared subunit of the TRRAP/TIP60 histone acetyltransferase and SRCAP complexes. J Biol Chem 2005; 280:13665 - 70; http://dx.doi.org/10.1074/jbc.M500001200; PMID: 15647280
- Wu WH, Alami S, Luk E, Wu CH, Sen S, Mizuguchi G, et al. Swc2 is a widely conserved H2AZ-binding module essential for ATP-dependent histone exchange. Nat Struct Mol Biol 2005; 12:1064 - 71; http://dx.doi.org/10.1038/nsmb1023; PMID: 16299513
- Jha S, Shibata E, Dutta A. Human Rvb1/Tip49 is required for the histone acetyltransferase activity of Tip60/NuA4 and for the downregulation of phosphorylation on H2AX after DNA damage. Mol Cell Biol 2008; 28:2690 - 700; http://dx.doi.org/10.1128/MCB.01983-07; PMID: 18285460
- Doyon Y, Selleck W, Lane WS, Tan S, Côté J. Structural and functional conservation of the NuA4 histone acetyltransferase complex from yeast to humans. Mol Cell Biol 2004; 24:1884 - 96; http://dx.doi.org/10.1128/MCB.24.5.1884-1896.2004; PMID: 14966270
- Smith ER, Eisen A, Gu W, Sattah M, Pannuti A, Zhou J, et al. ESA1 is a histone acetyltransferase that is essential for growth in yeast. Proc Natl Acad Sci U S A 1998; 95:3561 - 5; http://dx.doi.org/10.1073/pnas.95.7.3561; PMID: 9520405
- Sykes SM, Mellert HS, Holbert MA, Li K, Marmorstein R, Lane WS, et al. Acetylation of the p53 DNA-binding domain regulates apoptosis induction. Mol Cell 2006; 24:841 - 51; http://dx.doi.org/10.1016/j.molcel.2006.11.026; PMID: 17189187
- Tang Y, Luo J, Zhang W, Gu W. Tip60-dependent acetylation of p53 modulates the decision between cell-cycle arrest and apoptosis. Mol Cell 2006; 24:827 - 39; http://dx.doi.org/10.1016/j.molcel.2006.11.021; PMID: 17189186
- Bauer A, Huber O, Kemler R. Pontin52, an interaction partner of beta-catenin, binds to the TATA box binding protein. Proc Natl Acad Sci U S A 1998; 95:14787 - 92; http://dx.doi.org/10.1073/pnas.95.25.14787; PMID: 9843967
- Gospodinov A, Tsaneva I, Anachkova B. RAD51 foci formation in response to DNA damage is modulated by TIP49. Int J Biochem Cell Biol 2009; 41:925 - 33; http://dx.doi.org/10.1016/j.biocel.2008.09.004; PMID: 18834951
- Doyon Y, Côté J. The highly conserved and multifunctional NuA4 HAT complex. Curr Opin Genet Dev 2004; 14:147 - 54; http://dx.doi.org/10.1016/j.gde.2004.02.009; PMID: 15196461
- Volodin AA, Voloshin ON, Camerini-Otero RD. Homologous recombination and RecA protein: towards a new generation of tools for genome manipulations. Trends Biotechnol 2005; 23:97 - 102; http://dx.doi.org/10.1016/j.tibtech.2004.12.005; PMID: 15661347
- Cromie GA, Connelly JC, Leach DR. Recombination at double-strand breaks and DNA ends: conserved mechanisms from phage to humans. Mol Cell 2001; 8:1163 - 74; http://dx.doi.org/10.1016/S1097-2765(01)00419-1; PMID: 11779493
- McGlynn P, Lloyd RG. Action of RuvAB at replication fork structures. J Biol Chem 2001; 276:41938 - 44; http://dx.doi.org/10.1074/jbc.M107945200; PMID: 11551967
- Parsons CA, Tsaneva I, Lloyd RG, West SC. Interaction of Escherichia coli RuvA and RuvB proteins with synthetic Holliday junctions. Proc Natl Acad Sci U S A 1992; 89:5452 - 6; http://dx.doi.org/10.1073/pnas.89.12.5452; PMID: 1608954
- Hishida T, Han YW, Shibata T, Kubota Y, Ishino Y, Iwasaki H, et al. Role of the Escherichia coli RecQ DNA helicase in SOS signaling and genome stabilization at stalled replication forks. Genes Dev 2004; 18:1886 - 97; http://dx.doi.org/10.1101/gad.1223804; PMID: 15289460
- Fujiwara Y, Mayanagi K, Morikawa K. Functional significance of octameric RuvA for a branch migration complex from Thermus thermophilus. Biochem Biophys Res Commun 2008; 366:426 - 31; http://dx.doi.org/10.1016/j.bbrc.2007.11.149; PMID: 18068124
- Iwasaki H, Takahagi M, Shiba T, Nakata A, Shinagawa H. Escherichia coli RuvC protein is an endonuclease that resolves the Holliday structure. EMBO J 1991; 10:4381 - 9; PMID: 1661673
- Dunderdale HJ, Benson FE, Parsons CA, Sharples GJ, Lloyd RG, West SC. Formation and resolution of recombination intermediates by E. coli RecA and RuvC proteins. Nature 1991; 354:506 - 10; http://dx.doi.org/10.1038/354506a0; PMID: 1758493
- Shinagawa H, Iwasaki H. Processing the holliday junction in homologous recombination. Trends Biochem Sci 1996; 21:107 - 11; PMID: 8882584
- West SC. Processing of recombination intermediates by the RuvABC proteins. Annu Rev Genet 1997; 31:213 - 44; http://dx.doi.org/10.1146/annurev.genet.31.1.213; PMID: 9442895
- Shiba T, Iwasaki H, Nakata A, Shinagawa H. SOS-inducible DNA repair proteins, RuvA and RuvB, of Escherichia coli: functional interactions between RuvA and RuvB for ATP hydrolysis and renaturation of the cruciform structure in supercoiled DNA. Proc Natl Acad Sci U S A 1991; 88:8445 - 9; http://dx.doi.org/10.1073/pnas.88.19.8445; PMID: 1833759
- Hishida T, Han YW, Fujimoto S, Iwasaki H, Shinagawa H. Direct evidence that a conserved arginine in RuvB AAA+ ATPase acts as an allosteric effector for the ATPase activity of the adjacent subunit in a hexamer. Proc Natl Acad Sci U S A 2004; 101:9573 - 7; http://dx.doi.org/10.1073/pnas.0403584101; PMID: 15210950
- Michel B. Replication fork arrest and DNA recombination. Trends Biochem Sci 2000; 25:173 - 8; http://dx.doi.org/10.1016/S0968-0004(00)01560-7; PMID: 10754549
- Kolodner RD, Putnam CD, Myung K. Maintenance of genome stability in Saccharomyces cerevisiae. Science 2002; 297:552 - 7; http://dx.doi.org/10.1126/science.1075277; PMID: 12142524
- Branzei D, Foiani M. Template switching: from replication fork repair to genome rearrangements. Cell 2007; 131:1228 - 30; http://dx.doi.org/10.1016/j.cell.2007.12.007; PMID: 18160033
- Lambert S, Froget B, Carr AM. Arrested replication fork processing: interplay between checkpoints and recombination. DNA Repair (Amst) 2007; 6:1042 - 61; http://dx.doi.org/10.1016/j.dnarep.2007.02.024; PMID: 17412649
- Tourrière H, Pasero P. Maintenance of fork integrity at damaged DNA and natural pause sites. DNA Repair (Amst) 2007; 6:900 - 13; http://dx.doi.org/10.1016/j.dnarep.2007.02.004; PMID: 17379579
- Michel B, Boubakri H, Baharoglu Z, LeMasson M, Lestini R. Recombination proteins and rescue of arrested replication forks. DNA Repair (Amst) 2007; 6:967 - 80; http://dx.doi.org/10.1016/j.dnarep.2007.02.016; PMID: 17395553
- Baharoglu Z, Petranovic M, Flores MJ, Michel B. RuvAB is essential for replication forks reversal in certain replication mutants. EMBO J 2006; 25:596 - 604; http://dx.doi.org/10.1038/sj.emboj.7600941; PMID: 16424908
- Singleton MR, Dillingham MS, Gaudier M, Kowalczykowski SC, Wigley DB. Crystal structure of RecBCD enzyme reveals a machine for processing DNA breaks. Nature 2004; 432:187 - 93; http://dx.doi.org/10.1038/nature02988; PMID: 15538360
- Kiss T. SnoRNP biogenesis meets Pre-mRNA splicing. Mol Cell 2006; 23:775 - 6; http://dx.doi.org/10.1016/j.molcel.2006.08.023; PMID: 16973429
- King TH, Decatur WA, Bertrand E, Maxwell ES, Fournier MJ. A well-connected and conserved nucleoplasmic helicase is required for production of box C/D and H/ACA snoRNAs and localization of snoRNP proteins. Mol Cell Biol 2001; 21:7731 - 46; http://dx.doi.org/10.1128/MCB.21.22.7731-7746.2001; PMID: 11604509
- Gonzales FA, Zanchin NI, Luz JS, Oliveira CC. Characterization of Saccharomyces cerevisiae Nop17p, a novel Nop58p-interacting protein that is involved in Pre-rRNA processing. J Mol Biol 2005; 346:437 - 55; http://dx.doi.org/10.1016/j.jmb.2004.11.071; PMID: 15670595
- Zhao R, Kakihara Y, Gribun A, Huen J, Yang G, Khanna M, et al. Molecular chaperone Hsp90 stabilizes Pih1/Nop17 to maintain R2TP complex activity that regulates snoRNA accumulation. J Cell Biol 2008; 180:563 - 78; http://dx.doi.org/10.1083/jcb.200709061; PMID: 18268103
- Newman DR, Kuhn JF, Shanab GM, Maxwell ES. Box C/D snoRNA-associated proteins: two pairs of evolutionarily ancient proteins and possible links to replication and transcription. RNA 2000; 6:861 - 79; http://dx.doi.org/10.1017/S1355838200992446; PMID: 10864044
- McKeegan KS, Debieux CM, Boulon S, Bertrand E, Watkins NJ. A dynamic scaffold of pre-snoRNP factors facilitates human box C/D snoRNP assembly. Mol Cell Biol 2007; 27:6782 - 93; http://dx.doi.org/10.1128/MCB.01097-07; PMID: 17636026
- Boulon S, Marmier-Gourrier N, Pradet-Balade B, Wurth L, Verheggen C, Jády BE, et al. The Hsp90 chaperone controls the biogenesis of L7Ae RNPs through conserved machinery. J Cell Biol 2008; 180:579 - 95; http://dx.doi.org/10.1083/jcb.200708110; PMID: 18268104
- McKeegan KS, Debieux CM, Watkins NJ. Evidence that the AAA+ proteins TIP48 and TIP49 bridge interactions between 15.5K and the related NOP56 and NOP58 proteins during box C/D snoRNP biogenesis. Mol Cell Biol 2009; 29:4971 - 81; http://dx.doi.org/10.1128/MCB.00752-09; PMID: 19620283
- Cohen SB, Graham ME, Lovrecz GO, Bache N, Robinson PJ, Reddel RR. Protein composition of catalytically active human telomerase from immortal cells. Science 2007; 315:1850 - 3; http://dx.doi.org/10.1126/science.1138596; PMID: 17395830
- Kiss T. Small nucleolar RNAs: an abundant group of noncoding RNAs with diverse cellular functions. Cell 2002; 109:145 - 8; http://dx.doi.org/10.1016/S0092-8674(02)00718-3; PMID: 12007400
- Venteicher AS, Meng Z, Mason PJ, Veenstra TD, Artandi SE. Identification of ATPases pontin and reptin as telomerase components essential for holoenzyme assembly. Cell 2008; 132:945 - 57; http://dx.doi.org/10.1016/j.cell.2008.01.019; PMID: 18358808
- Matias PM, Gorynia S, Donner P, Carrondo MA. Crystal structure of the human AAA+ protein RuvBL1. J Biol Chem 2006; 281:38918 - 29; http://dx.doi.org/10.1074/jbc.M605625200; PMID: 17060327