Abstract
This essay provides a critical review of two recent books on evolution: Richard Dawkins’ The Greatest Show on Earth, and Jerry Coyne’s Why Evolution is True, as well as a critique of mainstream evolutionary theory and of natural selection. I also suggest a generalization of sexual selection theory that acknowledges mind as pervasive in nature. Natural selection, as the primary theory of how biological change occurs, must be carefully framed to avoid the long-standing “tautology problem” and must also be modified to more explicitly include the role of mind in evolution. A propensity approach to natural selection, in which “expected fitness” is utilized rather than “fitness,” can save natural selection from tautology. But to be a productive theory, natural selection theory should be placed alongside sexual selection – which is explicitly agentic/intentional—as a twin force, but also placed alongside purely endogenous factors such as genetic drift. This framing is contrary to the normal convention that often groups all of these factors under the rubric of “natural selection.” I suggest some approaches for improving modern evolutionary theory, including a “generalized sexual selection,” or "generalized agentic selection," a panpsychist extension of Darwin’s theory of sexual selection that explicitly recognizes the role of mind at all levels of nature and which may play the part of a general theory of evolution better than natural selection theory.
Background
The war of words between creationists and evolutionists goes on. But a middle way is becoming apparent – a theory of evolution that is neither dogmatically adaptationist nor dogmatically creationist. This middle way readily acknowledges that all life has evolved from common ancestors – a key difference from creationists. But the middle way does not necessarily accept that natural selection, defined narrowly as blind adaptation to environmental conditions, is the primary force driving evolution. Rather, consciousness (mind/agency) at various levels of nature is seen as a key force in evolution, perhaps the key force, along with endogenous factors such as drift and “laws of form.”
Neo-Darwinists, who are often “adaptationists” in that they assert that all or almost all evolution occurs through beneficial adaptations resulting from random variation, have upped the ante in the ongoing war in recent years with a number of books making the case for evolution and natural selection. The present essay is partly a review of two of these books, Richard Dawkins’ The Greatest Show on Earth,Citation1 and Jerry Coyne’s Why Evolution Is True,Citation2 as well as my own critique of mainstream evolutionary theory and a discussion of alternatives to neo-Darwinism.
Both of these books were published in 2009, the 150th anniversary of the publication of Darwin’s Origin of Species, the most influential book in biology. Dawkins (emeritus Oxford University, England) and Coyne (University of Chicago) are both highly respected biologists who have contributed greatly to their profession as well as to the public understanding of biology and science more generally. Dawkins, in particular, is known for his popular books explaining various aspects of evolution for lay audiences.
Dawkins and Coyne both attempt to create a new scientific terminology in their recent books. My feeling is that their suggestions in this regard are not helpful. They are fighting a long war with creationists and Intelligent Design proponents and have apparently decided to resort to extreme means in winning the war. It seems that they have become so frustrated over the years that their opponents will not simply accept their facts and theories on evolution as the gospel truth that they have abandoned scientific precision with respect to discussion of proof, fact, and theory.
Both Coyne and Dawkins suggest a new vocabulary and scientific methodology for discussing evolution. For Coyne, “evolution is a fact, natural selection is a fact.” I don’t find this revamping of scientific terminology helpful. Rather, a “fact” is a statement about the state of things at the time of the utterance, a statement about what is actual and objective – not a set of ideas about how things came to be as they are now. That’s rightly called a “theory.”Citation1 Facts are snapshots and theories are about how facts came to be. Theories are all about processes, change.
So evolution, defined simply as change in the biological world, is certainly a fact. We can see biological change happen in the test tube and in the natural world around us all the time, as well as in our own families from generation to generation. We see it happen with every sexual recombination, with every mutation, etc. Biological change is a constant fact. We simply compare snapshot to snapshot in the biological world and see constant change. Natural selection, however, is better described as a theory because it is not a statement about things as they are now compared with the most recent snapshots. It is, rather, a statement about how things came to be as they are now, over a long period of time (many many snapshots) – and about how species will evolve in the future. In other words, theories are about causes, which must always, under a Popperian philosophy of science, be inferred and never proven; facts are about observations.
Describing theories as “facts,” as Coyne does, is, it seems to me, a dangerous move designed to end debate. All theories are debatable because we can’t in science ever prove anything; we can only disprove things. Some theories certainly achieve dominance (the Modern Synthesis of evolutionary theory, for example), and some theories become truly hegemonic, but they should never be considered beyond debate.
Dawkins is better at discussing the “evidence for evolution” (the subtitle of his book but also the topic of Coyne’s book), whereas Coyne is better at discussing the theory of natural selection. Coyne devotes one chapter to natural selection, Dawkins not even that, an astounding omission in a book about mainstream evolutionary theory. (Climbing Mount Improbable,Citation3 an earlier Dawkins book, does discuss natural selection in detail). Dawkins states, as Coyne does, that evolution is a fact. Dawkins does not, however, state that natural selection is a fact. Rather, “it could still (just) be doubted that natural selection is [evolution’s] driving force.” (2009, p. 17). In other words, the evidence for evolution itself is so overwhelming that we should call this a fact, not a theory, but the evidence for natural selection as evolution’s driving force is not so strong as to call it a fact. It seems to me, however, as just discussed, that natural selection should never be called a fact, no matter how much evidence is asserted in its support, because natural selection is a theory about facts, not a fact itself.
My point is mirrored by Wilson, 1978,Citation4 (emphasis added):
Evolution – is it a fact or a theory? This question echoes an old and virulent controversy. It is important not only historically but also because of the light it can still shed on the distinction scientists make between fact and theory.
The process of evolution is a fact. It occurs. Biologists have watched and measured its progress at the level of the gene. They have created new species in the laboratory and in the experimental garden. They have collected a very large amount of fossil evidence, in many cases so complete that it cannot be rationally explained by any nonevolutionary hypothesis. On the other hand, how evolution occurs is a complex matter subject to theory.
Natural Selection as an Assumption
Coyne goes further than Dawkins and lumps natural selection with common descent as “evolution.” For Coyne, “evolution” means that natural selection is the cause of biological change, and not merely the fact of common descent – a more limited and traditional definition of evolution. Coyne’s approach doesn’t seem helpful to me because a growing number of biologists are concluding that natural selection may not be the most important or even a significant force in evolution in all circumstances. Rather, other forces, such as genetic drift (particularly in small populations), or other endogenous factors (collectively “internal selection”), Thompson (1961)Citation5 and Kauffman’s (1993)Citation6 “laws of form” and “order for free,”Citation2 the Baldwin Effect, the neo-Baldwin Effect, sexual selection, genetic assimilation, genetic accommodation, etc., may be more important in many circumstances. I describe at the end of this essay how sexual selection may be re-framed as a more general theory of evolution than the theory of natural selection. This re-framing is based on the recognition that mind is ubiquitous in nature, and thus choices, including most importantly mating choices, can apply in some manner to all entities.
Some biologists and philosophers (Karl Popper (1972),Citation7 most notably, though he later recanted) have concluded that natural selection is not a fact or a theory – it is, instead, a metaphysical assumption or “metaphysical research program” because it is unfalsifiable. Natural selection is, in this framing, the assumption that evolution occurs through natural forces instead of supernatural forces. Darwin wrote in The Descent of Man,Citation8 his second major work on evolution (p. 2, my emphasis):
I may be permitted to say, as some excuse, that I had two distinct objects in view [in writing The Origin of Species]; firstly, to shew that species had not been separately created, and secondly, that natural selection had been the chief agent of change ... ... hence if I have erred in giving to natural selection great power ... or in having exaggerated its power, which is in itself probable, I have at least, as I hope, done good service in aiding to overthrow the dogma of separate creations.
This assumption of natural causes for evolution is obviously extremely important in the history of evolutionary theory because it allowed us to proceed beyond an uninquisitive and dogmatic view of life as created and governed entirely by an active Creator God. But assumptions are not facts or theories. Assumptions are assumptions. That is, they are not necessarily supported by evidence and they are generally not provable or falsifiable within the theory they inform. They are the starting point for theories, not theories themselves.
For natural selection to avoid reducing to merely an assumption of natural forces behind evolution (rather than supernatural forces), we must carefully define natural selection as a theory of how evolution occurs, not a force or agent “selecting” traits, organisms or populations. This is particularly important because of the view that natural selection reduces to a tautology – a more threatening charge than that it reduces to merely an assumption of natural causes behind evolution. The next section discusses the tautology charge in detail, followed by a discussion of how the tautology problem can be avoided.
Natural Selection as a Tautology
Some thinkers have not been even so charitable as to call natural selection merely a metaphysical assumption, preferring instead to label it a tautology. Tautologies are true by definition. Tautologies are never false; therefore they are not falsifiable. Thus a tautology cannot be part of any empirical theory if we accept that empirical theories should be falsifiable.Citation3 Tautologies have the form of A = B, C, D… but reduce to A = A in actuality. That is, what seem to be different concepts (B, C, D …) all reduce in practice to A. Tautology in biology arises from the use of different terms for the same concept. A common way of describing natural selection is “survival of the fittest.” This phrase was coined by Herbert Spencer, a British philosopher, but was used by Darwin himself in later editions of Origin as a synonym for natural selection. Let’s look at the content of this phrase and its meaning.
Survival of the fittest means that the fittest organisms survive and thus spread more offspring. But we must ask ourselves what these terms mean. What does “the fittest” mean? Well, to be “fit” in this context means that those organisms that manage to survive leave more offspring. But what does “survival” mean? It means the same thing because there is no evolution without reproduction. So it turns out that the phrase “survival of the fittest” really means “survival of those who survive,” or “the fittest are the fittest.” “Survival of the fittest” states that A = B but in actuality the phrase reduces to A = A. A = A is always true, cannot be falsified, and is meaningless.
The statement, “natural selection is the process by which species change through survival of the fittest” is, then, a tautology with no explanatory (retrodictive) power and no predictive power because it is logically empty. It is akin to saying “evolution happens.” While this is obviously true, based on the abundant fossil record showing the development of life on our planet, it does not amount to a theory of how or why evolution happens. And the theory of natural selection should be that.Citation4
This is not a new critique of natural selection. In fact, Samuel Butler (1879),Citation9 a well-known critic of Darwin who had an ongoing feud with Darwin while both were alive, made this claim.Citation5 Many others have made the same argument since, including prominent biologists T.H. Morgan, C.H.Waddington,Citation6 and philosopher of science Popper, as well as more recently Rosenberg (2010),Citation10 a biologist and philosopher at Duke University. What follows are a few other examples for describing/defining natural selection, all of which also reduce to tautology.
I’ll show later that we can still define natural selection non-tautologically, but we have to be careful about it.
Natural selection is often described as “differential reproduction” of those organisms that have more adaptive traits. This means merely that some organisms leave more offspring than others (“differential”). If we assert something like “differential reproduction or survival of the fittest was responsible for the observed evolution,” we are, however, back to tautology because “differential reproduction,” “survival of the fittest” and “observed evolution” reduce to the same thing.
Another way of describing natural selection is by discussing “adaptive traits” or “adaptation” more generally. A trait is adaptive if it helps an organism survive and produce more offspring. But this is also tautological because the only way we have to determine what is adaptive is to examine an organism’s reproductive success. How else could we know what is adaptive? To say that a trait is adaptive and thus leads to more offspring is to say that a trait that leads to more offspring is a trait that leads to more offspring. We are back to tautology.
Gould (1976)Citation11 argued, as a solution to the tautology problem, for the discoverability of “a priori” engineering principles of adaptation that would provide reliable predictions for specific organisms and populations. These engineering principles, if they can in fact be discovered, would constitute the “meat” of the theory of natural selection. It remains to be seen what these a priori engineering principles would be and it seems that this approach is doomed to failure due to the same concerns already expressed: how would such principles be discovered other than by tracking what traits lead in real-world situations to improved survival and reproduction? Laboratory simulations, mechanical or virtual, can certainly help to develop hypotheses. But to know what the engineering principles of biology are in actuality, in the real world, requires that we observe what organisms survive and reproduce in real populations. There seems to be no other way. So Gould’s suggestion of discovering a priori engineering principles does not help us.
Here is the root of the problem. All these terms: survival, fitness, adaptation, differential reproduction, are referring to exactly the same concept – increased offspring and the spread of certain traits. Thus, to say that “natural selection is adaptation of organisms to their environment through survival of the fittest” sounds like it means something, but all this phrase really says is that increased offspring are increased offspring are increased offspring.
This is not only a problem that afflicts older descriptions of natural selection. It remains pervasive in how evolutionary theory is discussed. If evolutionary theory is to eventually dispel non-scientific rivals among the public, we who are on the side of science would be well-served in changing how we discuss natural selection and to rid it of tautological language. Here are a number of examples of tautological statements about natural selection from Coyne’s and Dawkins’ books:
Coyne states (ref. Citation2, p. 116): “Natural selection, we find, is everywhere, scrutinizing individuals, culling the unfit and promoting the genes of the fitter. It can create intricate adaptations, sometimes in surprisingly little time.” All of these phrases used by Coyne, “natural selection,” “scrutinizing individuals,” “culling the unfit,” “promoting the genes of the fitter,” reduce to exactly the same thing: biological change. They are different words for exactly the same outcome and provide no explanation at all of how or why any particular outcome occurred.
Coyne states (ref. Citation2, p. 117): “Natural selection, acting on coat color [in oldfield mice], has simply changed the genetic composition of a population, increasing the proportion of genetic variants … that enhance survival and reproduction.” Again, “natural selection,” “changed the genetic composition,” “increasing the proportion of genetic variants,” and “that enhance survival and reproduction” all reduce to exactly the same thing: biological change has occurred.
Coyne states (ref. Citation2, p. 117): “[Natural selection] is not a mechanism imposed on a population from outside. Rather, it is a process, a description of how genes that produce better adaptations become more frequent over time.” Again, “natural selection,” “process,” “better adaptations,” and “become more frequent over time” reduce to the same thing.
Dawkins states (ref. Citation1, p. 352): “But most mutations are disadvantageous, if only because they are random and there are many more ways of getting worse than there are ways of getting better. Natural selection promptly penalizes the bad mutations. Individuals possessing them are more likely to die and less likely to reproduce, and this automatically removes the mutations from the gene pool.” “Natural selection,” “promptly penalizes,” “more likely to die and less likely to reproduce,” and “automatically removes the mutations from the gene pool” are all phrases representing the same concept. A = A = A = A.
Dawkins has provided a more concise definition of natural selection (from another of his books, Climbing Mount Improbable, which Coyne quotes approvingly): Natural selection is the “non-random survival of random variants.” This is also tautological, however, because “non-random” and “survival” reduce to each other. What is non-random survival? The only way we can know is by looking at those that survive. Which ones survive? The non-randomly surviving ones. In this case “non-random” is a stand-in for “the fittest.” It tells us nothing more about which individuals or populations are likely to survive in any real-world situation (or lab situation, for that matter) than saying “those that survive survive.” Alternatively, we can merely note that saying survival is “non-random” tells us nothing in itself about what organisms or populations are likely to survive in actual circumstances, leaving the theory empty in a different way. (Dawkins updates this definition in The Greatest Show on Earth (p. 405), without resolving the clear tautology: Natural selection is “the non-random survival of information that encodes embryological recipes for that survival.”)
The Propensity Interpretation of Fitness
All is not lost. There is a way out of the tautology problem if we replace “fitness” with “expected fitness” and make explicit the fact that there is no actual quality or propensity, labeled fitness or expected fitness, inhering in organisms or populations. Expected fitness is purely a theoretical tool that saves natural selection from tautology and allows for potentially useful predictions and retrodictions. If we realize that fitness or, more accurately, expected fitness is just a theoretical tool and consists of nothing a priori (as Gould argued) we may avoid tautology and reification of concepts as dispositions that inhere in organisms and populations.
The “propensity interpretation” of natural selection was developed to better explain the concept of fitness, by replacing “fitness” with “expected fitness,” and is defined by Beatty and Finsen (1989)Citation12 as follows: “fitter organisms have a greater ability to leave offspring, and not just (and not necessarily) greater success in this regard.” This approach does allow for a non-tautological interpretation of natural selection,Citation7 by focusing on the “ability” (propensity or disposition) to leave offspring rather than actual offspring; but this path has its own problems. Beatty and Finsen conclude: “We are not sure whether a radically revised interpretation of fitness is necessary. But it does seem to us that certain gross oversimplifications of the propensity interpretation deserve more serious attention. We most certainly do not propose to return to the interpretation of fitness that the propensity interpretation was designed to replace. Whatever fitness is, it is not actual offspring contribution, although it was long misconceived as such.”
“Whatever fitness is…” is the key phrase here. Beatty and Finsen undertake a detailed examination of the many different propensity interpretations, which they label the “multiple propensities problem,” demonstrating that “the greater ability to leave offspring” is itself impossible to define in a non-arbitrary way. This is the case because what timeframe should one consider in such an analysis? One generation, two, three, infinite? And how do we measure offspring? Different statistical approaches yield different answers. Beatty and Finsen illustrate this problem with the following image: is A or B more fit; does A or B have a higher propensity for survival and thus a higher “expected fitness” ()?
Figure 1. Illustration of Beatty and Finsen’s “multiple propensities problem”.
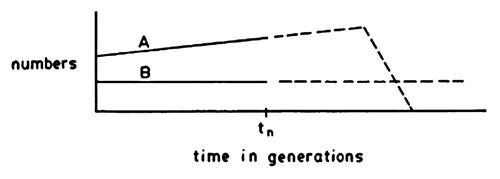
The inescapable conclusion (my own, not Beatty and Finsen’s, who hold out some hope of a non-arbitrary approach) is that “expected fitness” is neither a disposition nor ability and it cannot be defined in a non-arbitrary way – but this isn’t itself a fatal problem if we recognize that “expected fitness” is simply the biologist’s tool for making predictions or retrodictions about expected evolution in actual organisms or populations (which I’ll flesh out further later).
Rosenberg and Bouchard (2008)Citation13 introduce the many difficulties facing the concept of “fitness” in their Stanford Encyclopedia of Philosophy entry on “fitness”:
But what is fitness and how can one tell when a trait enhances fitness, or more to the point, when one organism is fitter than another? Opponents of the theory of natural selection have long claimed that the theory is so treated by its proponents as to define fitness in terms of rates of reproduction, thus condemning the principle of the survival of the fittest to triviality: the claim that those organisms with higher rates of reproduction leave more offspring is an empty, unfalsifiable tautology bereft of explanatory power. In the century and a half since the publication of On the Origin of Species biologists have all too often reinforced this objection by actually so defining fitness. For example, C.H. Waddington writes, in Towards a Theoretical Biology, the fittest individuals are those that are “most effective in leaving gametes to the next generation.” It appears therefore that evolutionary theory requires a definition of fitness that will protect it from the charges of tautology, triviality, unfalsifiabilty, and consequent explanatory infirmity. If no such definition is in fact forthcoming, then what is required by the theory's adherents is an alternative account of its structure and content or its role in the research program of biology.
They add: “It is no wonder that biologists … have defined ‘x is fitter than y’ in terms of quantitatively measurable reproductive rates. This tendency of course simply adds force to the original argument [that fitness is a tautological concept]. If the only way to make fitness-differences scientifically tractable is to trivialize the theory, so much the worse for the theory.”
With respect to the propensity interpretation of fitness, Rosenberg and Bouchard state that evolutionary theory’s “reliance on the concept of ‘fitness’ makes it imperative that conceptual problems threatening the explanatory legitimacy of this notion be solved.” And: “[F]ar from providing the theoretical meaning of fitness, the probabilistic propensity ‘definition’ is a set of an indefinitely large number of operational measures of fitness.” They conclude: “The problem of defining fitness remains.”
There is No Process of Natural Selection; There is a Theory of Natural Selection
I’ve shown that no matter how we describe natural selection as a process that takes place in nature such descriptions either reduce to the assumption of natural forces rather than supernatural forces behind natural selection, and not a theory of evolution in itself. Or we arrive at tautology.Citation8 There is thus no merit in talking about natural selection as a process – even this statement is arguably tautological because “natural selection” and “process” reduce to the same thing in this context (A = A). If we define natural selection, however, as a theory that attempts to explain biological change through the expected fitness (propensity for more offspring, which is necessarily arbitrarily defined) of each organism or population, rather than the actual fitness, we are saved from tautology but we are left without a general theory of evolution. To be clear, I am suggesting that natural selection should be defined as “the theory that attempts to explain biological change through the expected fitness of each organism.” I suggest this approach because setting “expected fitness” as the dependent variable does not itself constitute a theory at all. This approach still leaves blank the crucial factors relevant to the expected fitness (“expected fitness of a given population is a result of …”; or “A, expected fitness, is the result of B, C, D, etc.”). In other words, for any actual circumstance, what are the factors that will guide the biologist’s prediction of expected fitness? These factors require additional theoretical structure.
The various factors thought to be relevant to evolutionary change must be examined and included in the prediction of expected fitness. Again, this is not a fatal problem for the propensity interpretation and it is perhaps the only way out of the tautology problem. I’ll propose some approaches for filling in the blanks in the theory, building on the propensity interpretation of fitness, in the last section of this essay.
Adaptationism vs. Pluralism
Whereas Darwin’s vision was “pluralist” because he suggested many agents for evolution (natural selection, sexual selection, habit, etc.), today’s mainstream evolutionary theory is generally “adaptationist” in that it invokes natural selection as either the only significant cause of evolution (adaptation) or at least its primary agent. “Genetic drift” and many other agents are also recognized by mainstream biology but the large majority of biologists still stress natural selection as the key agent. Adaptationists see all, or almost all, evolution resulting from natural selection acting on random variation of different traits. Dawkins states, for example, in The Greatest Show on Earth (p. 332), without a denial, “I have a reputation as an arch-‘adaptationist.’”
There is a healthy debate today between adaptationists and pluralists, but this debate is unfortunately confined to a small subset of biologists and philosophers of biology. Steven Orzack politely demolishes the “messy state of the art” in testing natural selection and adaptationism in the 2010 Oxford Handbook of the Philosophy of Biology.Citation14 Orzack concludes that only three (literally) sets of studies from the last two centuries of biology were sufficiently rigorous to count as tests of “optimality models” with respect to natural selection. Optimality models are constructed to make predictions about expected evolutionary change in populations as a result of natural selection (re-framed as “optimality” because it is suggested that natural selection will in most cases result in optimal adaptations for a given environment). All other optimality tests, according to Orzack, either simply assume that natural selection is the primary or only mechanism of change, without further consideration – begging the question in the extreme – or are insufficiently quantitative to establish anything significant with respect to optimality for the traits at issue. More generally, Orzack concludes (p. 87):
[E]volutionary biologists often use models having assumptions known to be manifestly false or at best of uncertain validity in order to predict evolutionary outcomes. It’s a conceptual and logical mess, and consequently, one might be tempted to regard the enterprise as dubious at best. Nonetheless, many such attempts at prediction are remarkably successful.
A broader issue with adaptationism is that its root concept – “adaptation” – is even more problematic than “fitness.” One is hard pressed to find a definition of adaptation in the literature but the generally accepted meaning of the term is “a trait that enhances the ability of an organism to survive or reproduce.” But, as discussed above, how can we know that any particular trait enhances survival of an organism or population unless we conduct tests that distinguish particular traits from others? This is the root of the long-standing debate over “spandrels” and “free-riders.” Gould and Lewontin (1979)Citation15 analogized biological free-riders with architectural spandrels in European cathedrals. Spandrels are the triangular areas between arches, which result necessarily from joining two or more arches to support a dome. The spandrel is, thus, a free-rider on the architect’s choice to use arches. The analogy is meant to suggest that in biology we often cannot distinguish arches from spandrels in that we often cannot distinguish what was selected-for, because it was adaptive, from what is merely selected, but not selected-for, as a spandrel/free-rider. That is, the biological free-rider is an accidental concomitant of selection-for, of adaptation.
A recent example is illustrative. The New York TimesCitation16 contained an insightful article on recent evolutionary theory, stating:
Dr. Barrett [one of the biologists focused on in the article] studied the evolution of the three-spine stickleback, a three-inch-long fish that lives both in oceans and in freshwater lakes. In the oceans, the fish are covered with armor. But in lakes, they are virtually armor-free.
There was an “adaptive story” about the sticklebacks, as there was with the deer mice. The same armor that had protected the ocean sticklebacks from larger fish proved deadly in the lakes, making them easier for predatory insects to catch. So over time, they shed the armor.
But that story turned out to be flawed. In fact, the armor gene was linked to a gene for rapid growth, which gave the lake fish an advantage by allowing them to breed earlier and better survive cold winters. Growth rate, not armor, was determining survival — and driving evolution.
This is the kind of issue that led Gould and Lewontin to write their 1979 “Spandrels paper” already cited – they warned against the too-frequent habit of biologists to consider plausible scenarios about how organisms could have evolved to be sufficiently explanatory with respect to how they really did evolve. But because it is often very difficult to distinguish free-riders from selected-for traits, these explanations often amount to “just so” stories about evolution.Citation9 In order to establish the validity of adaptationism, Orzack has called for an Adaptationism Project akin to the Human Genome Project. This project would conduct and collect rigorous studies from around the world and establish in as many real circumstances as possible whether natural selection was at work, or was the primary agent in observed evolutionary change.
Other Critiques of Coyne and Dawkins
Another key step in resolving the problems identified is recognizing the role (perhaps the starring role) of mind in evolution. Coyne and Dawkins stress in their books the “mindless” nature of evolution by natural selection, as do most biologists today. But this is demonstrably wrong – even under their own terms. Both authors discuss sexual selection at some length, considered generally by modern biologists to be a sub-set of natural selection, and both acknowledge that sexual selection is largely about mating choice (generally female choice). The choices made by females drive sexual selection; just as human mating choices require a mind, so such choices require minds in non-human animals. Evolution is thus, even for these committed materialist Darwinians, imbued with mind at every level of at least the animal kingdom.
For example, Coyne describes sexual selection as a subset of natural selection, which is a common view today, but then insists that in natural selection: “There is no will involved, no conscious striving.” (ref. Citation2, p. 117). What is male-male competition in mating competitions and in pursuing mates if not will, if not conscious striving? What is female choice in mate selection if not will? Even if we restrict our consideration to sexual selection in humans, it is quite obvious that will and conscious striving are very much involved. We don’t have to be Lamarckians to realize that conscious striving is ubiquitous at least in the animal kingdom in terms of the struggle to survive and reproduce.
My next critique, applicable to both books, is also a compliment. Both do a great job of providing evidence for common descent and both tell engrossing stories about evolving whales, rabbits, humans, etc. This is because these stories are highly interesting in themselves, but also, and more to the point, they comprise the key evidence that Coyne and Dawkins want to convey in order to combat the Creationist story of special creation (which is directly opposed to the idea of common descent). While there are unfortunately legions of people in the US and elsewhere who still do not believe in common descent, the far more interesting questions to those of us trained in biology and science is not whether evolution has occurred, but how evolution has occurred and how it will work in the future. Natural selection is the dominant theory of evolutionCitation10 today and yet neither book spends much time on this key theory (Coyne far more than Dawkins).
An Alternative View
There are alternatives to natural selection and adaptationism as theories of evolution, including a set of concepts developed by D’Arcy Wentworth Thompson, a 20th Century British biologist, and Stuart Kauffman, a contemporary American biologist, that focus on the “laws of form” (ref. Citation6) and “order for free” (ref. Citation7) in nature as a whole. Rather than looking to natural selection as the key agent of change in evolution, these thinkers see order arising spontaneously in all sorts of places around us, such as in snowflakes, crystals, as well as in extremely complex organisms like ourselves and many other creatures, as a compounding of these more basic sources of order. In other words, much of biological change can be explained through the same chemical and physical principles that hold at simpler levels of nature.
A little ironically, Dawkins states that Thompson’s book “is not a book on evolution,” but about ontogenetic development, contrary to Thompson’s obvious intent. The quote in footnote 1 of the present essay, in which Thompson describes all of nature (physical, chemical, biological), as “so many riddles of form, so many problems of morphology,” makes it clear that Thompson was indeed concerned about evolution as well as development because we cannot have development without evolution. They are, in fact, two aspects of exactly the same process: the progression of life, moment to moment, year to year, from its origin to the present.
Another, more controversial, notion is that there is a driving force behind complexity and evolution. This idea has been championed by various biologists, philosophers and theologians over the centuries, starting with Aristotle’s entelechy. LamarckCitation17 was one of the early and most prominent thinkers in the modern era who suggested that evolution was being pushed by a “force that perpetually tends to make order.” This was one of two agents of evolution that Lamarck proposed. The second is more well-known and considered discredited today: the notion that organisms themselves, through intention, use and disuse, can change their bodies and that some of these changes are inherited. There is, however, an increasing body of evidence that some evolutionary change is Lamarckian. C. H. Waddington, Ted Steele, Eva Jablonka, Marion Lamb, Mary Jane West-Eberhard and others have argued in recent years for at least partial acceptance of Lamarckian inheritance of acquired traits, arguing that while it is obvious that not all acquired traits are inherited, some are. I will explore these ideas further below.
The final concept I’ll mention was described well by the American philosopher Gerald Heard. Heard wrote in his classic 1939 essay, Pain, Sex and Time,Citation18 that “from the most primitive forms of life up to the completion of man’s physique, the one clear coordinating achievement is heightened awareness.” Modern biologists know that there is no necessary progression from lower complexity and awareness to higher complexity and awareness in every circumstance. We have many examples of organisms becoming less complex as they evolve. However, it is undeniable that there is a general trend, as Heard describes, toward greater complexity and greater awareness (perception), which far outweighs the anomalous examples of reduction in biological complexity. Evolution is generally synonymous with complexification and heightened perception/awareness.
There is some interesting recent scholarship in the area of “cognitive biology” that is relevant to my arguments, with perhaps Kovac’sCitation19 work most prominent. Kovac writes in a 2008 paper on bioenergetics and the human brain: “The reason why cognition has become the most accelerating factor of evolution is straightforward: the growth of knowledge, noogenesis, is autocatalytic, and hence exponential or even hyperbolic.”Citation20
Recognizing that this trend toward greater awareness and increased cognition exists, we can propose as a working hypothesis that there is a cause for this trend. This process may reduce to the same “order for free” tendency that Thompson and Kauffman focus on, compounded at many levels over time. But Heard suggests, and I agree, that there is more going on here than the simple physical and chemical ordering principles that Thompson and Kauffman focus onCitation11 Rather, there seems to be a basic force in all things that leads to greater connection, thus greater complexity, and thus greater awareness of our universe around us. No intelligent designer needs to be postulated, nor mystical additional forces, as an explanation here, but there does seem to be a process occurring in the evolution of life on our planet that can’t be explained through the traditional (materialist) concepts of mindless matter and energy – or is at least difficult to explain from a traditional materialist perspective. Rather, it seems that we can better explain the observed trends through a re-framing away from the materialist conception of matter and an increased emphasis on individual (organismic) selection, in which each individual is the source of much evolutionary change.
Modern evolutionary theory is framed in terms of the evolution of populations. What is often missed, however, is that evolution of populations cannot happen without the evolution of individuals. Evolution in populations is simply the collective evolution of individuals. My addition to this picture is an increased emphasis on the role of individuals, each with minds, in shaping their own evolution – and thus the evolution of populations.
Alfred North Whitehead, a British mathematician, physicist and philosopher who ended his long career at Harvard, as professor of philosophy from 1924 to 1937, argued in his later works that all matter, literally, has some degree of experience, of awareness. This view is known as panpsychism or panexperientialism and is an increasingly popular solutionCitation12 to the broad inquiry over the last few decades into the nature of consciousness. Panpsychism’s increasing popularity has occurred in part because of the difficulties facing the “emergentist” concept of consciousness, in which consciousness, it is suggested, springs forth from wholly mindless matter at some point in the course of evolution and development (phylogenetically and ontogenetically). As the American biologist Sewall Wright (1977)Citation21 succinctly states: “Emergence of mind from no-mind is sheer magic.” In other words, positing that mind arises from that which is defined as no-mind faces very serious philosophical problems. Wright finds it incredible, as do I, which requires that we think more broadly about the nature of mind/consciousness in relation to matter.
It turns out that panpsychism offers not only a powerful solution to the question of “what is consciousness?” but also to the question of “how did life arise and evolve?” The panpsychist solution is to recognize that mind and thus purpose are inherent in all of nature – but are extremely rudimentary in most cases. However, as matter complexifies in molecules like amino acids (which form spontaneously in many situations), this innate mind and purpose starts to play an increasingly significant role in evolution. It is, thus, a bootstrapping process that has no end in sight.
Margulis and Sagan, two respected but admittedly non-mainstream contemporary biologists, support this view in their highly readable 1995 book, What is Life?Citation22 They appeal to Samuel Butler, an early critic of Darwin’s (p. 232):
Butler brought consciousness back in [to biology] by claiming that, together, so much free will, so much behavior becoming habit, so much engagement of matter in the processes of life, had shaped life, over eons producing visible organisms, including the colonies of cells called human. Power and sentience propagate as organisms. Butler's god is imperfect, dispersed. We find Butler's view - which rejects any single, universal architect - appealing. Life is too shoddy a production, both physically and morally, to have been designed by a flawless Master. And yet life is more impressive and less predictable than any 'thing' whose nature can be accounted for solely by 'forces' acting deterministically.
In evolution, then, God is indeed in the details – literally. The “dispersed” God that Margulis and Sagan refer to is the mind contained in each thing, in each organism, that exercises some degree of choice – no matter how small – in how it manifests. This is the Generalized Sexual Selection (GSS) I describe below, which is an elaboration of Darwin’s own ideas on sexual selection. In GSS, all things have male and female aspects, and (primarily female) choice has played a strong role in the evolution of life on our planet; perhaps the starring role. When we combine Thompson and Kauffman’s “order for free” with the panpsychist Generalized Sexual Selection, we may arrive at a general theory of evolution that can complement or perhaps even rival natural selection as a more complete theory for explaining evolution.
Generalized Sexual Selection
Darwin’s second major work, the 1871 Descent of Man and Selection in Relation to Sex, focused in large part on sexual selection, another agent of evolution that he realized was theoretically necessary to explain features like the peacock’s tail (Darwin stated in a letter to Asa Grey: “The sight of a peacock's tail, whenever I gaze at it, makes me sick!”). Why is sex so central to our lives? The facile answer is that it’s because we need sex to reproduce. But this is only partly true. Many species reproduce without sex, including some complex vertebrates like lizards and fish. So why do we have sex? No one really knows the answer to this question, but there are many theories. I won’t delve much into why our species reproduces sexually; rather, I’m going to delve into what sex is, as a general principle, and the role of sex in evolution.
Sexual selection is the term Darwin gave to the idea that certain traits appear to be detrimental to survival and/or foraging for food – such as the peacock’s tail, yet if such traits help an organism find more mates and have more offspring the trait may still spread. This is because the trait’s benefits in terms of reproduction outweigh its disadvantages in terms of survival. Mate choice, primarily female choice because males are generally the aggressors in most sexually-reproducing species, is key to sexual selection. Sexual selection has two key components: mate choice and male-male competition. There are many other aspects to sexual selection, however, including: mutual mate choice, assortative mating, male selection on females, which are all important in socially monogamous birds, prosimians, humans, and some other species.
Sexual selection is “agentic” in that it is based on intentions associated with minds. So whereas natural selection – or “survival selection,” as it is perhaps more accurately described – is explicitly non-agentic, as stressed by its proponents like Coyne and Dawkins, sexual selection relies on minds as agents of evolution. However, “agentic selection” (as Miller uses this term in The Mating Mind: How Sexual Choice Shaped the Evolution of Human Behavior)Citation23 is a bridging concept that is broader than sexual selection as Darwin conceived of it. This is the case because there are many types of agentic selection that don’t directly involve sex or reproduction. (My theory could probably be called “Generalized Agentic Selection” without losing much. I prefer, however, Generalized Sexual Selection, because of the recognized history of sexual selection as a theory.)
See for various types of agentic and sexual selection. It’s important to keep in mind that “sexual selection” in this table refers to the traditional notion of sexual selection and not the expanded “generalized sexual selection” that I describe further below ().
Table 1. Aspects of sexual selection and agentic selection.
Sexual selection may provide the basis for a more general theory of evolution than natural selection. Historically, these two selective “forces” have been presented by biologists as parallel forces, but with natural selection as by far the more important force. In recent decades, sexual selection is generally presented as a sub-set of natural selection, though it seems to me that sexual selection should be considered a parallel force that is perhaps in many cases more powerful than the environmental influences of natural selection. Alternatively, sexual selection, particularly in the generalized form I focus on here, may act as the content of natural selection theory, which, as discussed above, reduces to an assumption that evolutionary change occurs through natural and not supernatural forces.
In reality, of course, there is no “force” behind natural selection as it is generally framed. It’s all just physics and chemistry in action; so when biologists talk about natural selection as a force or an agent, it’s reification at work; there is no force producing change. Sexual selection is different, however, because there really is supposed to be a selective agent (a force of a sort) at work, which may not be explained wholly through physical and chemical forces – if these forces and their theoretical structure ignore mind in nature, as they generally do in the prevailing materialist paradigm. Contemplating these ideas requires that we consider whether mind (and thus choices made by minds) can in fact be explained through current physical and chemical theories. This is where evolutionary biology and the philosophy of mind intersect.
i. I argued in Hunt (2011), “Kicking the Psychophysical Laws into Gear: A New Approach to the Combination Problem,”Citation24 that current physical theories cannot, in principle, explain mind because the constituents of matter are defined by modern physics as wholly mindless. Mind is, under prevailing physical theories, described as an emergent phenomenon of certain kinds of complex biological structures like humans; but the emergence of mind from what is defined as wholly mindless requires a miracle (as Wright states). We are thus left with a system of physics that excludes that which is most real to each of us – ourselves, our own minds, subjectivity itself – which surely should be included in an adequate theory of physics and, by extension, biology. I argued that this impasse requires the inclusion of mind, in at least a highly rudimentary form, in all forms of matter from the simplest to the most complex. This view is known as panpsychism, as discussed above.
This discussion raises the question: how did we, and other life forms like us, reach such a high level of complexity? How did we evolve?
If we acknowledge that all matter has at least some rudimentary type of mind, no matter how simple, we realize that some degree of choice must be inherent in all matter. At its most basic level, mind requires perception and a choice. This is the case because mind may be defined as the selection (choice) between alternatives made available through perception of potential futures. We can’t know what an electron’s experience is like, but I presume that it consists of little more than rudimentary perceptions of the outside world, through its being influenced by the fundamental forces of electromagnetism, gravity, etc., and a choice as to how to manifest in the next moment based on those rudimentary perceptions (recall that each subject oscillates between object and subject, requiring its re-creation in each and every moment). Indeed, for Whitehead choice is the key feature of actuality: “’[D]ecision’ cannot be construed as a casual adjunct of an actual entity. It constitutes the very meaning of actuality.”Citation25
This notion is similar to Schopenhauer’s concept of the “will”Citation26 as fundamental to reality, though with a less negative connotation than it held for that gloomy philosopher. The physicist Freeman Dyson (Professor Emeritus at Princeton’s Institute for Advanced Study) makes my point explicit with respect to subatomic particlesCitation27: “[T]he processes of human consciousness differ only in degree but not in kind from the processes of choice between quantum states which we call ‘chance’ when made by electrons.”Citation13 This notion is very strange to those encountering it for the first time, but there are many arguments in its favor, which I flesh out in Hunt 2011.
Dyson is saying that what physicists normally interpret in electron behavior as pure chance – randomness – is better interpreted as choice. Choice not chance. Choices can be fickle, so what seems to be random is better interpreted as a result of unpredictable choices by these tiny entities. Thus, even electrons make choices – but very simple choices compared with the infinity of choices possible to our advanced human consciousness. Choice at the level of the electron is apparently limited to where the electron will manifest in the next moment. Particles such as electrons are not static, timeless entities. Thinking of the fundamental constituents of reality as unchanging particles is the fallacy of “substantialism,” which Whitehead’s panpsychist “process philosophy,” attempts to correct.
If choice is inherent even at the level of electrons, and every level above and below, a universal principle of evolution is made apparent. I call this universal principle “generalized sexual selection.” The essence of sexual selection is choice – generally female choice, as Darwin described in The Descent of Man. Darwin argued that female choice resulted over many generations in pronounced features in males who compete vigorously for female attention.
The simplest structure of neo-Darwinian natural selection has just two parts: 1) random variation of traits results from random mutation of genes and through sexual recombination; 2) some traits spread because they confer a reproductive advantage.
What I call generalized sexual selection (GSS or “giss”), re-frames this argument as follows: 1) variation in traits comes about through random mutation and through male competition for mating opportunities and striving more generally for self-improvement, which can sometimes be incorporated into the germ line of the male. In other words, variation is not always random – it is sometimes directed,Citation14 with improved mating opportunities as a significant motivation. The urge to survive, and other urges also, are surely at work in prompting this non-random source of variation.Citation15 2) Natural selection, as the sum total of environmental influences, molds organisms and populations, but (generally) female choice is often a powerful selective agent that leads to greater reproduction of those males with the most desirable traits for the females who choose them, who incorporate the male germ line into their own by mating with them. Thus, selection is not blind, it is conscious at every level of nature through the choices made by each entity – when we recognize that consciousness exists in at least a rudimentary manner at every level of nature, an idea that I discuss further below. GSS may fairly be described as “neo-Lamarckian” because individual striving (choice) plays a large role in evolution.
Perhaps the most clear-cut examples of Lamarckian inheritance today involve the immune system. See Steele, et al.Citation28 for an extended argument that retrogenes (proteins impact RNA and DNA, contrary to the normal view) are the rule in the immune system. A very interesting recent example of Lamarckian inheritance involves the green sea slug, which can incorporate algae genes into its soma, live off sunlight alone, and pass this ability on to its offspring.Citation29 I'm not suggesting that the slug has any high-level awareness of what it is doing, but it is certainly striving to eat the algae, with all the consequences thereof ensuing. More generally, Nowacki, Landweber, et al.,Citation30 of Princeton continue to study Lamarckian paths of inheritance in ciliates and other creatures. Experiments have established that ciliates often display Lamarckian inheritance. BussCitation31 takes a rigorous look at all high-level taxa and catalogs which ones demonstrate Lamarckian inheritance and which don't (Buss 1983 finds that fungi, protists and plants all demonstrate Lamarckian inheritance in some circumstances, and this kind of inheritance is also present in a number of animal phyla).
A new field, behavioral epigenetics, is essentially Lamarckian in that it recognizes explicitly that the Central Dogma (soma cannot influence DNA) is invalid. David Sweatt, at the University of Alabama, Binghamton, stated in a 2011 article: “There’s no dichotomy between genes and environment. Instead there is a constant dynamic interplay between genome and environment.”
Moshe Szyf, a pharmacologist at McGill University who was one of the first to relate DNA methylation patterns to behavior, sees epigenetic processes as “the adaptive mechanism of the genome”—an essential mechanism for pruning down the wide range of all possible behaviors permitted by genes, selecting those that fit an individual’s environment. “DNA methylation is a physiological mechanism,” he says, “by which the genome senses the world and changes itself.”
West-Eberhard (2003)Citation32 presents a wealth of information and argument with respect to phenotypic plasticity resulting from “genetic accommodation,” a relatively new term for what may be considered a neo-Lamarckian mechanism of evolution.Citation16 Her work was the first effort to systematically describe the new field of “evo-devo,” evolutionary developmental biology. Her work is somewhat dated because of the rapid growth in this field, but she nevertheless paints a comprehensive picture of the role of the environment and learning on variation and selection. Similar to Waddington’s “genetic assimilation,” genetic accommodation is environmentally induced evolution (genetic assimilation is a type of genetic accommodation), which includes organismic learningCitation17 and thus a role for mind and choice. There is still a role for random mutation, but West-Eberhard suggests that it plays a weaker role than the environment and intention of the organism at issue. She summarizes her thesis (p. vii): “The universal environmental responsiveness of organisms, alongside genes, influences individual development and organic evolution, and this realization compels us to reexamine the major themes of evolutionary biology in a new light.”
In sum, it seems clear that the tendency since the late 19th Century to dismiss Lamarckian mechanisms, as either a priori impossible or merely rare,Citation18 was premature and unwarranted.
With respect to selection, I suggest that sexual selection should be recognized as a more powerful force for evolution than is generally acknowledged and that mind and intention are the key to this evolutionary process. Sexual selection has made something of a comeback after being all but ignored for many decades after Darwin first proposed it in 1871. Cronin (1993, p. 4)Citation33 states:
But to a Darwinian, [characteristics such as the peacock’s tail or the eusocial behavior of ants] pose a common difficulty. Wouldn’t natural selection be expected to eliminate, rather than favour, them? For over a century these problems, when not neglected, were 'solved' in quite erroneous ways… But in the last few decades Darwinism has undergone a revolutionary change. And, in the wake of this transformation, the obstinate anomalies of altruism and sexual selection are anomalies no more.
An interesting new area of research has sprung up since Cronin’s book that further bolsters the place of sexual selection in evolutionary theory. Weiner (1995)Citation34 presents a highly readable account of American biologists Peter and Rosemary Grant’s work on Galapagos finches over a 2year period. The Grants, well-known now for this work, came to realize that hybrid breeding – that is, cross breeding of different species of finches on the same islands – has at times played a very strong role in the evolution of these birds. Grant, et al. (2003, p. 2915, citations omitted)Citation35 concludes: “Occasional hybridization may have important evolutionary potential … in complex ways that are still being explored ….” Grant and Grant (2010),Citation36 a genetic analysis of hybridization among the same populations of finches, concludes:
The relatively large populations of Geospiza fortis and G. scandens [two of the finch species the Grants studied] receive conspecific immigrants at a rate of less than one per generation. They exchange genes more frequently by rare but repeated hybridization. Effects of heterospecific gene flow from hybridization are not counteracted by lower fitness of the offspring. As a result, the standing genetic variation of the two main resident populations on Daphne Major is enhanced to a greater extent by introgressive hybridization than through breeding with conspecific immigrants.
Kaneshiro has explored similar themes in his work on Hawaiian Drosophila, arguing that sexual selection and the sub-set of sexual selection known as hybridization has played a larger role than generally thought in speciation. Hybridization is an interesting case of sexual selection because it suggests that sexual attraction, when broad enough to ensure some breeding with individuals not normally part of the same breeding group, may be a powerful force for evolutionary change.
My thesis is, however, that additional modification is required for the theory of sexual selection to be fully realized, and the key change is to recognize the role of mind throughout nature. I call my proposed approach “generalized sexual selection” because all evolutionary forces may be framed as types of sexual selection – if we recognize that sex goes beyond what we generally think of when we use this term. As very sexual primates, we Homo sapiens see sex as key to reproduction and much of human interaction. However, most species on our planet do not reproduce sexually in the traditional sense. Bacteria, for example, often reproduce asexually, as do protists. And even many vertebrates reproduce asexually, such as certain species of lizards and fish. This definition is, however, overly restrictive and fails to indicate that almost all species do indeed exchange genetic information at some point during reproduction. Bacteria are constantly exchanging genetic information, which is a rudimentary kind of sex, if we define sex at this level, in an expanded but entirely legitimate manner, as the mixing of genetic information from at least two entities. This type of sex, in my expanded definition, is generally known as “horizontal gene transfer” because it occurs without simultaneous reproduction.
But GSS applies even further beyond the traditional and my expanded definition of sex. The terms “male” and “female” are not as clear-cut as we generally assume. In GSS, “male” refers to any genetic donor and “female” to any genetic recipient – as Margulis and Sagan describe in their 1986 book, The Origins of Sex. Thus, a bacterium that gives some genetic material to another is a male and the recipient is a female. These roles can and do change on a regular basis, thus the “gender” of each bacterium changes regularly. What is important, then, is not gender, per se, but actions.
This principle extends even deeper, however, when we consider further the panpsychist notion of matter. If all matter is associated to some degree with mind or subjectivity, then GSS applies to literally all matter, not just biological forms. This is the case because the ultimate constituents of matter – what Whitehead calls “actual entities” – contain both “mental” and “physical” aspects. They are two sides of the same coin. Physical and mental aspects of each actual entity (the Whiteheadian “atom”) oscillate with each step forward in time. Matter is not, in Whitehead’s system, comprised of static entities jostling against each other in deterministic patterns through the eons, but is, instead, comprised of events that are forever changing non-deterministically. Each event/actual entity interacts with countless other events in the process that produces actuality from potentiality in each moment.
The mental aspect of each actual entity is informed by the immediately prior physical aspects of all other actual entities available to it as the source of data/perceptions. Each actual entity, in its mental aspect, chooses what information to accept and rejects everything else. Thus, the mental aspect of each actual entity can be considered to be “female” insofar as it chooses what information from the universe around it to include in its objective manifestation – like the female bower bird accepting the attention of a hard-working showy male. When the actual entity becomes objective, it becomes “male” insofar as its manifestation now constitutes data for the next iteration of actual entities to consider in their mental/female aspect. More crudely put, the female aspect receives and the male aspect penetrates. But these aspects oscillate within each actual entity at simpler levels of nature and only become generally fixed as more complex biological forms appear.
GSS is a re-framing of evolution in a way that recognizes the unbroken continuum of the complexity of matter, which is experiential through and through, from the simplest of structures to the most complex. I won’t delve into further details about the testability and falsifiability of GSS here, but it is my view that GSS presents a more adequate theory of evolution than the prevailing adaptationist view of natural selection – which generally denies the role of mind and choice in evolution. compares three approaches to evolutionary theory.
Table 2. Comparing theories of evolution
My suggested alternative to adaptationist natural selection, GSS, may be summarized as follows, described more fully in HuntCitation37 (2012, in preparation):
• Fitness does not exist and nor does “expected fitness” in an ontological sense; rather, expected fitness is merely a theoretical tool for making evolutionary predictions, not a disposition or quality that inheres in organisms or populations.
• Variation in traits results from random endogenous processes but also from conscious striving and habit, which probably become more important as organisms become more complex, resulting in an increasing pace of evolution.
• Trait variation in ontogeny and phylogeny is both limited and guided by Thompson and Kauffman’s laws of form/order for free.
• Propagation of traits occurs through a combination of (inorganic) environmental factors, interspecies competition, intra-species competition for resources and mates, and (generally) female mating choices (the latter two factors are generally described as “sexual selection”). These factors are collectively described as “selection factors.”
• Agentic evolution, as opposed to non-agentic evolution (purely environmental selection), is highlighted in sexual selection and to a lesser degree in interspecies competition.
• Hybridization is increasingly recognized as a major factor in evolution and speciation – and hybridization is a type of sexual selection that occurs when different individuals cross the prevailing attraction barriers (which define a species or variety) to mate with one another.
• All selection factors may more fundamentally be described as variations of my proposed generalized sexual selection theory.
The benefits of this approach are many: we achieve new clarity in our terminology and resolve contradictions in existing evolutionary theory (such as classifying sexual selection as a subset of natural selection, which makes little sense because they often work at cross-purposes); we recognize mind in nature as a powerful force for change at all levels, providing content to evolutionary theory beyond the principle/assumption of natural selection, which stands only for the view that natural causes lead to evolutionary change; and we help explain how genetic variation can lead to beneficial traits by recognizing that variation is not all random, particularly at higher levels of complexity. Rather, a highly important source of variation is induced by individuals striving to improve their survival or their mating prospects.
Time will tell if this approach catches on, but it seems clear that modern evolutionary theory must evolve to incorporate mind explicitly in its explanations if we are to better understand nature in all its grandeur. Coyne and Dawkins have certainly helped sway some readers from the dogma of creationism, but the far more interesting debate is not between creationism and Darwinism – it is between materialism and scientific alternatives to materialism such as panpsychism.
The “endless forms most beautiful” that Darwin revered are indeed beautiful but also endlessly fascinating. These forms are made even more fascinating by the realization that what is most real to each of us – our own consciousness, our subjectivity – is most likely equally real for all levels of nature and has played a key role in producing the ongoing complexification of life.
Notes:
a Dawkins suggests a less harmful, but I think equally unhelpful, neologism: he suggests we should discuss evolution as a “theorum,” meant to suggest that evolution itself has the (almost) certainty of a mathematical theorem – not the colloquial meaning of “theory,” which often suggests to the layperson that an idea is half-baked and “just a theory.”
b D’arcy Thompson, the British polymath, wrote in his magnum opus, On Growth and Form (1917, 1961): “The waves of the sea, the little ripples on the shore, the sweeping curve of the sandy bay between the headlines, the outline of the hills, the shape of the clouds, all these are so many riddles of form, so many problems of morphology… Nor is it otherwise with the material forms of living things. Cell and tissue, shell and bone, leaf and flower, are so many portions of mater, and it is in obedience to the laws of physics that their particles have been moved, moulded and conformed.”
c It was because of this lack of falsifiability of the theory of natural selection that Popper preferred to label the theory a “metaphysical research program,” which is still highly valid as part of science but according to Popper shouldn’t be considered a gold standard scientific theory.
d Lewontin (1970) writes: “The logical skeleton of [Darwin’s] argument … turns out to be a powerful predictive system for changes at all levels of biological organization.”
e “The fact that one in a brood or litter is born fitter for the conditions of its existence than its brothers and sisters, and, again, the causes that have led to this one’s having been born fitter—which last is what the older evolutionists justly dwelt upon as the most interesting consideration in connection with the whole subject—are more noteworthy factors of modification than the factor that an animal, if born fitter for its conditions, will commonly survive longer in the struggle for existence. If the first of these can be explained in such a manner as to be accepted as true, or highly probable, we have a substantial gain to our knowledge. The second is little—if at all—better than a truism. Granted, if it were not generally the case that those forms are most likely to survive which are best fitted for the conditions of their existence, no adaptation of form to conditions of existence could ever have come about. “The survival of the fittest” therefore, or, perhaps better, “the fertility of the fittest,” is thus a sine quâ non for modification.”
f “The general principle of natural selection, in fact, merely amounts to the statement that the individuals which leave the most offspring are those which leave the most offspring. It is a tautology.” C.H. Waddington in The Strategy of the Genes.
g Brandon and Ramsey (2007) state: The prime motivation [for introducing the propensity interpretation] was to make room for an explanatory theory of natural selection, which is tantamount to solving the so-called “tautology problem.” This problem arises from a casual inspection of the phrase “survival of the fittest” and then asking what defines the fittest. If the answer is those that reproduce the most, then it seems we are explaining a phenomenon, differential reproduction, in terms of itself, which is no explanation at all.”
h Scientific theories are sometimes described as tautological as a general matter and this is sometimes offered as a defense to the tautology problem in evolutionary theory. For example, in physics force equals mass times acceleration (f = ma, which is Newton’s Second Law of Motion) and this is a rather useful equation. This is not, however, a tautology because it is falsifiable. We could perform experiments and find that force does not equal mass times acceleration, falsifying this law of physics. Newton’s Second Law is in fact generally considered to already have been falsified through Einstein’s expansion of Newtonian physics with the theories of special and general relativity. This defense thus offers no help for those combating the tautology problem in evolution.
i Fodor and Piatelli-Palmerini (2010) argue forcefully that the “free-rider problem” is fatal to natural selection as a theory because the empirical investigations required to establish what is selected-for rather than a free-rider collectively constitute “natural history.” It is, then, natural history, not “natural selection,” that can in particular cases explain evolution. As such, the theory of natural selection is not explanatory of anything. This is another way of arriving at the conclusion that natural selection reduces to an assumption that evolution proceeds through natural forces and not supernatural forces, as discussed earlier in this essay. These authors make a strong argument, but it seems unfair to dismiss natural selection as a theory because it requires fleshing out in particular cases. As I’ve discussed, I think we can frame natural selection as a productive theory, if we’re careful, but I also argue later in this essay that an alternative “generalized sexual selection” theory may offer a more general and explanatory theory of evolution.
j Hopi Hoekstra, professor and curator of mammals for the Harvard Museum of Comparative Zoology, stated in a 2011 New York Times article (Rosner 2011): “Fitness is the most important concept in biology …. But no one ever measures it.” This isn’t entirely true but it is the case that it is too little measured or considered critically as a concept central to biology.
k Thompson himself states in On Growth and Form: “My sole purpose is to correlate with mathematical statement and physical law certain of the simpler outward phenomena of organic growth and structure or form, while all the while regarding the fabric of the organism, ex hypothesi, as a material and mechanical configuration. This is my purpose here. But I would not for the world be thought to believe that this is the only story which Life and her Children have to tell. One does not come by studying living things for a lifetime to suppose that physics and chemistry can account for them all.” (P. 9).
l Many respected thinkers have subscribed to some form of panpsychism, including the ancient Greeks Heraclitus and Empedocles, Plotinus in the 3rd Century CE, to Giordano Bruno in the 16th Century, Spinoza and Leibniz in the 17th Century, Immanuel Kant (in his earlier work) in the 18th Century, Arthur Schopenhauer and Ernst Haeckel in the 19th, and in the 20th Century William James, Gregory Bateson, the biologists J.B.S. Haldane, Sewall Wright, and C.H. Waddington, the paleontologist and theologian Pierre Teilhard de Chardin, the physicists Sir Arthur Eddington, Whitehead, David Bohm, Freeman Dyson, Roger Penrose, and the philosopher Bertrand Russell (to some degree). More recently, Galen Strawson, Stuart Hameroff, David Chalmers, William Seager, Gregg Rosenberg, Jonathan SchoolerCitation1 and many others have advocated panpsychist or quasi-panpsychist views. The debate has intensified also in Germany, which has long been more receptive to panpsychism than the US - see e.g., the new book, Müller T, Watzka H . Ein Universum voller »Geistestaub«?: Der Panpsychismus in der aktuellen Geist-Gehirn-Debatte. 2011.
m A similar point is made in Seager W. (1995) Consciousness, Information and Panpsychism, Journal of Consciousness Studies, 2, 3, pp. 272–88, and Bohm D. and Hiley B. (1993) The Undivided Universe: An Ontological Interpretation of Quantum Theory, Oxon: Routledge.
n Rossi (Rossi, Ernest Lawrence. The Psychobiology of Gene Expression: Neuroscience and Neurogenesis in Hypnosis and the Healing Arts. New York: W.W. Norton &, 2002. Print) explores in detail how mental states can induce rapid genetic changes, such as Immediate Early Gene expression in humans, which can become active within a matter of seconds as a response to emotional states. Rossi describes a continuum of genetic expression triggered by the environment and mental states, overturning the Central Dogma of modern evolutionary theory, which holds that information only flows from genes to proteins and not the other way.
o Kirschner and Gerhart (Kirschner, Marc W., and John C. Gerhart. The Plausibility of Life. New Haven.: Yale UP, 2005. Print) provide additional explanatory power with respect to the non-randomness of variation, but I don’t have space in this brief essay to delve into their ideas and demonstrate how their ideas are complementary to GSS.
p West-Eberhard disavows any Lamarckian taint (“There is no hint of direct (Lamarkian [sic]) influence of environment on genome in this scheme…,” p. 29), but when we consider West-Eberhard’s other statements in the same book and other work, these concepts are certainly neo-Lamarckian in the key sense that there is some degree of inheritance of acquired traits. She states, for example (p. 115): “In fact, given continuity of the phenotype, transmission of acquired phenotypes can be considered a property of all ontogenies.” It seems that West-Eberhard’s discomfort with the label “Lamarkian” stems from concern about avoiding the controversy this label brings. She writes on the same page: “The unhappy fate of processes labeled Lamarkian is that they are likely to be labeled as oddities even if common and important.” This aversion may be wise, and time will tell, but I believe there is little value in avoiding certain labels, when accurate otherwise, merely because of previous controversies.
q “Extreme plasticity such as learning can produce exceedingly rapid (abrupt) speciation.” (P. 526).
r Maynard Smith (1989) states: “Lamarkian inheritance is rare.”
Related Research Data
References
- Dawkins R. The Greatest Show on Earth: the Evidence for Evolution. London: Black Swan, 2010.
- Coyne JA. Why Evolution Is True. New York: Viking, 2009.
- Dawkins R. Climbing Mount Improbable. New York: Norton, 1996.
- Wilson EO. Life on Earth. Sunderland, MA: Sinauer Associates, 1978.
- Thompson D'arcy W. On Growth and Form: an Abridged Edition. Cambridge: Cambridge UP, 1961.
- Kauffman SA. The Origins of Order: Self-organization and Selection in Evolution. New York: Oxford UP, 1993.
- Popper KR. Objective Knowledge; an Evolutionary Approach. Oxford: Clarendon, 1972.
- Darwin C. The Descent of Man and Selection in Relation to Sex. 2nd edition. Detroit: Gale Research, 1974.
- Butler S. Evolution, Old and New; Or, the Theories of Buffon, Dr. Erasmus Darwin, and Lamarck, as Compared with That of Mr. Charles Darwin. London: Hardwicke and Bogue, 1879.
- Rosenberg A, Bouchard F. “Fitness (Stanford Encyclopedia of Philosophy).” Stanford Encyclopedia of Philosophy. Web. 17 Dec. 2011.
- Gould SJ. “Stephen Jay Gould, “Darwin's Untimely Burial” 1976.” Harper's Weekly (1976). The Unofficial Stephen Jay Gould Archive. Web. 17 Dec. 2011.
- Beatty J, Finsen S. Rethinking the Propensity Interpretation of Fitness: A Peak Inside Pandora's Box. In: Ruse M. What the Philosophy of Biology Is: Essays Dedicated to David Hull. Springer, 1989.
- Rosenberg A, Bouchard F. “Fitness (Stanford Encyclopedia of Philosophy).” Stanford Encyclopedia of Philosophy. Web. 17 Dec. 2011.
- Orzack SH. Testing Adaptive Hypotheses, Optimality Models, and Adaptationism. In: Ruse M. Oxford Handbook of Philosophy of Biology. Oxford: Oxford UP, 2010.
- Gould SJ, Lewontin R. The spandrels of San Marco and the panglossian paradigm: a critique of the adaptationist programme. Proc R Soc Lond B 205 1979; 1161:581-98.
- Rosner H. A Colorful Way to Watch Evolution in Nebraska’s Sand Dunes. New York Times [New York] 8 Aug. 2011.
- Lamarck JB. Philosophie Zoologique, Ou, Exposition, Des Considérations Relatives À L'histoire. Bruxelles: Culture Et Civilisation, 1983.
- Heard G. Pain, Sex and Time; a New Outlook on Evolution and the Future of Man. New York and London: Harper & Brothers, 1939.
- Kovác L. Bioenergetics: A key to brain and mind. Commun Integr Biol 2008; 1(1):114-22; Jedlicka P: Physical complexity and cognitive evolution, In: Gershenson C, Aerts D, Edmonds B (eds), Worldviews, science and us, World Scientific Publishing, Singapore, pp. 221-23; Kovác L: Information and knowledge in biology: time for reappraisal. Plant Signal Behav 2007 Mar; 2(2):65-73.
- Kovác L. Bioenergetics: A key to brain and mind. Commun Integr Biol 2008; 1:114 - 22; http://dx.doi.org/10.4161/cib.1.1.6670; PMID: 19513208
- Wright S. Panpsychism and Science. In: Cobb JB, Griffin DR (eds). Mind in Nature: Essays on the Interface of Science and Philosophy. Lanham, MD: University Press of America, 1977.
- Margulis L, Sagan D. What Is Life? New York: Simon & Schuster, 1995.
- Miller G. The Mating Mind: How Sexual Choice Shaped the Evolution of Human Behavior. New York: Anchor, 2001.
- Hunt T. Kicking the psychophysical laws into gear: a new approach to the combination problem. J Conscious Stud 2011; 18:96 - 134
- Whitehead AN. Process and Reality. New York, NY: The Free Press, 1929. P. 43.
- Schopenhauer A. The World as Will and Representation. New York: Everyman, 1995.
- Dyson FJ. Disturbing the Universe. New York: Harper & Row, 1979.
- Steele EJ, Lindley RA, Blanden RV. Lamarck's Signature: How Retrogenes Are Changing Darwin's Natural Selection Paradigm. Reading, MA: Perseus, 1998.
- latimes.com/news/nationworld/nation/la-sci-sea-slugs16-2010jan16,0,861315.story.
- Nowacki M, Vijayan V, Zhou Y, Schotanus K, Doak TG, Landweber LF. RNA-mediated epigenetic programming of a genome-rearrangement pathway. Nature 2008; 451:153 - 8; http://dx.doi.org/10.1038/nature06452; PMID: 18046331
- Buss LW. Evolution, development, and the units of selection. Proc Natl Acad Sci U S A 1983; 80:1387 - 91; http://dx.doi.org/10.1073/pnas.80.5.1387; PMID: 6572396
- West-Eberhard MJ. Developmental Plasticity and Evolution. New York: Oxford UP, 2003.
- Cronin H. The Ant and the Peacok Altruism and Sexual Selection Form Darwin Today. Cambridge: Cambridge UP, 1993.
- Wiener J. The Beak of the Finch. London: Vintage, 1995.
- Grant PR, Grant BR, Keller LF, Markert JA, Petren K. Inbreeding and interbreeding in Darwin’s finches. Evolution 2003; 57:2911 - 6; PMID: 14761069
- Grant PR, Grant BR. Conspecific versus heterospecific gene exchange between populations of Darwin’s finches. Philos Trans R Soc Lond B Biol Sci 2010; 365:1065 - 76; http://dx.doi.org/10.1098/rstb.2009.0283; PMID: 20194169
- Hunt T. Kicking the psychophysical laws into gear, Part 2: Toward a general theory of biology. 2012, in progress.