
Abstract
Many see fruit flies as an annoyance, invading our homes with a nagging persistence and efficiency. Yet from a scientific perspective, these tiny animals are a wonder of multisensory integration, capable of tracking fragmented odor plumes amidst turbulent winds and constantly varying visual conditions. The peripheral olfactory, mechanosensory, and visual systems of the fruit fly, Drosophila melanogaster, have been studied in great detail;1-4 however, the mechanisms by which fly brains integrate information from multiple sensory modalities to facilitate robust odor tracking remain elusive. Our studies on olfactory orientation by flying flies reveal that these animals do not simply follow their "nose"; rather, fruit flies require mechanosensory and visual input to track odors in flight.5,6 Collectively, these results shed light on the neural circuits involved in odor localization by fruit flies in the wild and illuminate the elegant complexity underlying a behavior to which the annoyed and amazed are familiar.
Figures and Tables
Figure 1 Olfactory orientation in flight requires multisensory input. (A, top) In a free-yaw magnetic tether flight arena,Citation7 flying flies were positioned with an oscillating vertical stripe 90° to the right (blue arrows) of a 20° apple cider vinegar plume (orange triangle). At time = 0, we switched on a high contrast visual panorama and recorded the subsequent turning behavior by the fly (data for A-C adapted fromCitation5). (A, bottom) Five representative flight trajectories, where color is used to distinguish between individuals, reveal flies direct saccades toward the plume. Occluding the left 3rd antennal segment with glue (B, top) abolishes leftward olfactory orientation (B, bottom). Immobilizing the left Johnston's organ (JO) with glue (C, top) abolishes leftward olfactory orientation (C, bottom). When presented with a uniform visual panorama (D, top) at time = 0, instead of a high contrast visual panorama (A, top), flies fail to orient toward the odor plume (D, bottom). (experimental methods for D as in (A) and;Citation5 visual stimulus as inCitation6)
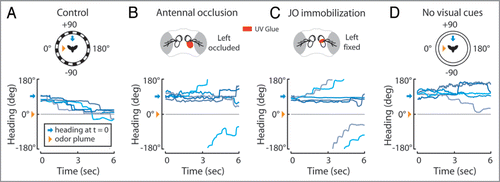
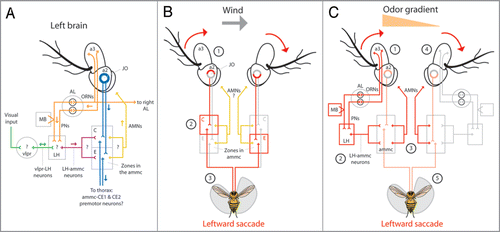
Figure 2 Anatomical model for olfactory orientation. (A) Odor activates first-order olfactory receptor neurons (ORNs) housed in the 3rd antennal segment (a3) which connect unilaterally, or bilaterally to specific second-order olfactory projection neurons (PNs) within defined glomeruli in the antennal lobeCitation9 (AL, orange). PNs connect to either the lateral horn (LH) or the mushroom body (MB) and the (LH).Citation11 The LH presumably receives visual input from the ventrolateral protocerebrum (vlpr) via vlpr-LH interneuronsCitation13 (green) and apparently relays visual and olfactory information to the antennal mechanosensory and motor center (ammc) via LH-ammc interneuronsCitation11,Citation13 (maroon). The ammc receives input from the mechanosensory Johnston’s organ (JO) housed in the second antennal segment (a2, blue). JO neurons project to defined stimulus specific regions of the ammc;Citation18 zones C and E mediate wind sensation.Citation1 Presumably, antennal motor neurons (AMNs, yellow) arise from unknown regions of the ammc and innervate muscles within the a2s to rotate the a3s. Rotation of a3 relative to a2 is encoded by JO neurons which may synapse onto identified pre-motor descending neurons; the dendrites of ammc-CE1 and ammc-CE2 descending neurons innervate zones C and E of the ammc.Citation3 (arrows indicate presumed direction of information flow; (?) indicate unknown connectivity) (B) Wind from the left passively rotates both a3s clockwise (1) and through JO neurons, activates zone C ipsilateral to the wind and zone E contralateral to the wind (2). Gluing the left JO impairs leftward upwind orientationCitation16 suggesting that premotor neurons connecting to zone C of the left ammc activate leftward saccades (3). It is unknown if AMNs participate in this response. (C) As a result of , we believe that the increased activation of the left ORNs and PNs in response to an odor on the left (1, orange triangle) and via LH-ammc neurons (2),Citation13 triggers an asymmetrical activation of AMNs arising from the ammc (3) which, via antennal muscles in the a2s,Citation21 actively rotate the a3s to mimic a passive wind stimulus (4) and trigger a leftward ammc mediated saccade (5, ). (red arrows in B and C indicate rotation of a3 relative to a2; red lines in B and C indicate presumed and exaggerated circuit activation; orange triangle in C indicates higher odor concentration on the left).
Addendum to: