
Abstract
Though recent work has demonstrated that plants can recognize species, kin versus strangers, and self/non-self roots, no mechanism for identity recognition in plants has yet been found. Here we examined the role of soluble chemicals in signaling among roots. Utilizing Arabidopsis thaliana, we exposed young seedlings to liquid media containing exudates from siblings, strangers (non-siblings), or only their own exudates. In one experiment, root secretions were inhibited by sodium orthovanadate and root length and number of lateral roots were measured. In a second experiment, responses to siblings, strangers, and their own exudates were measured for several accessions (genotypes), and the traits of length of the longest lateral root and hypocotyl length were also measured. The exposure of plants to the root exudates of strangers induced greater lateral root formation than exposure of plants to sibling exudates. Stranger recognition was abolished upon treatment with the secretion inhibitor. In one experiment, plants exposed to sibling or stranger exudates have shorter roots than plants only exposed to their own exudates. This self/non-self recognition response was not affected by the secretion inhibitor. The results demonstrate that that kin recognition and self/non-self are two separate identity recognition systems involving soluble chemicals. Kin recognition requires active secretion by roots.
Acknowledgments
H.P.B acknowledges the University of Delaware-EPSCoR for faculty start-up grant and NSF (0713774). S.A.D. acknowledges the Natural Science and Engineering Research Council of Canada for research funding. Authors also thank Drs. Thomas E. Hanson, Madhavi Avadhani and Nicole Donofrio for critical reading of the manuscript and editing suggestions, Kathleen Donohue and Robin Cameron for providing seed, and Sinah Lee and Delicia S. Moonilal for technical help. S.A.D. thanks Robin Cameron for her generous help with techniques and supplies.
Figures and Tables
Figure 1 Lateral roots in Arabidopsis thaliana plants grown solitary (OWN) or in sibling (KIN) and non-sibling (STRANGER) exudates from the secretion inhibitor experiment. (A) Shows the representative photographs of OWN, KIN and STRANGER seedlings in the control treatment. (B) Shows the representative photographs of OWN, KIN and STRANGER treatments in the sodium vanadate treatment. (C) Shows the average number of lateral roots among OWN, KIN and ortho STRANGER seedlings in control and sodium vanadate (Na3VO4 [3 µM]) treatments. Means with the same letter do not differ significantly. Ortho bars indicate mean± S.E. for N = 864.
![Figure 1 Lateral roots in Arabidopsis thaliana plants grown solitary (OWN) or in sibling (KIN) and non-sibling (STRANGER) exudates from the secretion inhibitor experiment. (A) Shows the representative photographs of OWN, KIN and STRANGER seedlings in the control treatment. (B) Shows the representative photographs of OWN, KIN and STRANGER treatments in the sodium vanadate treatment. (C) Shows the average number of lateral roots among OWN, KIN and ortho STRANGER seedlings in control and sodium vanadate (Na3VO4 [3 µM]) treatments. Means with the same letter do not differ significantly. Ortho bars indicate mean± S.E. for N = 864.](/cms/asset/8b651226-b2ce-4d43-a31a-07ef487cf718/kcib_a_10910118_f0001.gif)
Figure 2 Average length of primary roots in Arabidopsis thaliana plants grown solitary (OWN) or in sibling (KIN) and non-sibling (STRANGER) exudates from the secretion inhibitor experiment, for the control and sodium orthovanadate [Na3VO4 (3 µM)] treatments. Means with the same letter do not differ significantly. Bars indicate mean± S.E for N = 864.
![Figure 2 Average length of primary roots in Arabidopsis thaliana plants grown solitary (OWN) or in sibling (KIN) and non-sibling (STRANGER) exudates from the secretion inhibitor experiment, for the control and sodium orthovanadate [Na3VO4 (3 µM)] treatments. Means with the same letter do not differ significantly. Bars indicate mean± S.E for N = 864.](/cms/asset/246286bb-e9dd-40d1-9e5a-b87d3f21e00e/kcib_a_10910118_f0002.gif)
Figure 3 Average lateral root number for eight accessions of A. thaliana in OWN (solitary), KIN (from siblings), and STRANGER (non-siblings) exudates. Trial 1 comprises CHA 11, CHA 38, trial 2 Col(0) and WS(0), and trial 3 CHA 28, CHA 31, CHA 34, and CHA 4. Average lateral root number is greater in Col(0) and WS (0) accessions than CHA accessions, but the effects of exudates did not differ among accession. Bars indicate means ± 1S.E for N = 216.
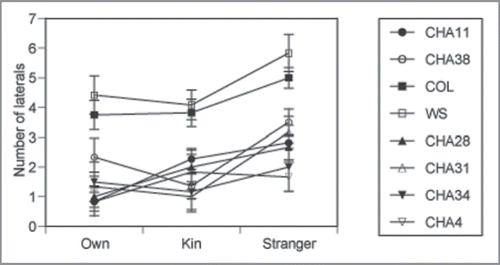
Figure 4 The panel A, B, C represents the experimental set up for OWN, KIN and STRANGER treatments. The arrows in the panel indicates the transfer of plants into exposure to KIN/STRANGER root exudates.
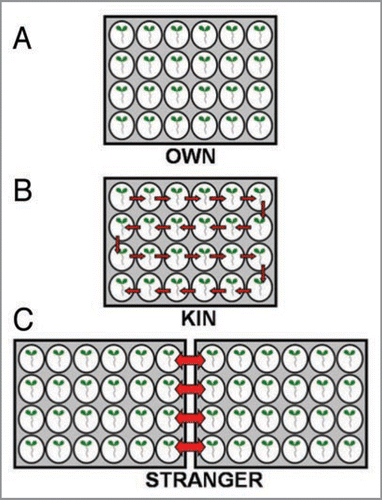
Table 1 Mixed model analysis of variance for the secretion inhibitor experiment. Fixed effects are exudate source (OWN, KIN and STRANGER), secretion inhibitor (control and sodium orthovanadate) and trial. Well plates are considered random effects nested within the three way interaction of exudate source, secretion inhibitor, and trial. Degrees of freedom for numerator and denominator, and F-ratios are reported for fixed effects, and z-values for random effects. For lateral roots, initial analysis showed that the three way interaction between kin, exudate inhibitor, and trial was non-significant with F4.18=0.29 and P> 0.8833. Because the limited degrees of freedom severely compromise the power of the analysis, we dropped that non-significant three-way interaction.
Table 2 Mixed model analysis of variance for the multiple accession experiments. Fixed effects are exudate source (OWN, KIN and STRANGER) and trial. Only one well plate was done for each treatment at each trial, so the trial × exudate source interaction is considered a random effect. Each accession was included in only one trial, so accession is nested within trial as a random effect, and the interaction of exudate source × genotype nested with trial is also a random effect. Degrees of freedom for numerator and denominator, and F-ratios are reported for fixed effects, and z-values for random effects.
Table 3 Pearson correlations among seedling traits from the multiple accession experiment.