
Abstract
The ability of an organism to detect a predator and then to take the appropriate vigilance actions is paramount for survival of the species. Lab-reared snails (> 250 generations) maintain their ability to detect predators and alter both aerial and cutaneous respiration. However, only the scent of a sympatric predator altered aerial respiration in freshly collected ‘wild’ snails. Here we test the hypothesis that the detection of a sympatric predator but not an allopatric predator will alter cutaneous respiration in freshly collected ‘wild’ snails. We find that Alberta snails while altering their cutaneous respiration to the scent of a sympatric predator (Tiger salamander) do not alter respiration to the scent of a crayfish (an allopatric predator). In Dutch snails there is a greater alteration to the scent of crayfish (sympatric predator) than to an allopatric predator (Tiger salamander).
Figures and Tables
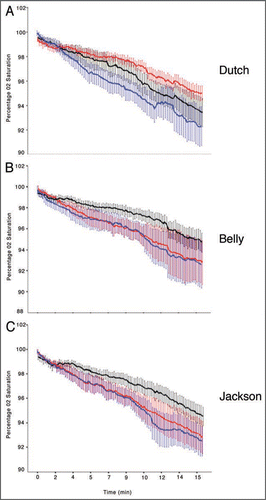
Figure 1 Non-aerial oxygen consumption in a closed chamber between three populations of Lymnaea stagnalis in PW, CE and SE. (A) O2 consumption of Dutch snails in three water treatments (N = 9, p < 0.01 for each pair-wise comparison). All three water treatments are significantly different from one another. The rate of O2 decrease in Dutch snails is lowest in CE (top red trace) then SE (middle black trace) and is highest in PW (bottom blue trace). (B) O2 consumption on Belly snails was significantly reduced in SE (N = 9, p < 0.01 for each pair-wise comparison, top black trace) but were not significantly different between PW and CE (N = 9, p > 0.05, bottom red and blue overlapping traces). (C) O2 consumption in Jackson snails is similar to that of Belly snails in that SE was significantly reduced compared to PW and CE (N = 9, p < 0.01 for each pair-wise comparison, top black trace) but were not significantly different between PW and CE (N = 9, p > 0.05, bottom red and blue overlapping traces).
Addendum to: