Abstract
It was recently discovered that the photoreceptor cryptochrome is involved in mediating magnetosensitive entrainment of the internal clock of fruit flies (1). This discovery follows other recent studies implicating a role of cryptochrome in mediating magnetic sensitivity in orientation responses of fruit flies (2,3) and growth responses of plants (4). Such widespread use of the same molecule for mediating magnetic sensitivity might suggest that cryptochrome is in some way optimal for detecting the magnetic field of the earth and that the magnetoreception function cannot be easily taken over by other molecules. This raises the question what properties might set cryptochromes apart from other molecules in terms of their ability to detect the geomagnetic field. Here, we will discuss possible answers to this question. We will first review the likely biophysical mechanism by which magnetic fields can be detected by a photoreceptor and discuss what constitutes an optimal photo-magneto-receptor. We will then discuss in how far cryptochrome matches the profile of an optimal molecule and what further steps are required for more conclusive answers.
Figures and Tables
Figure 1 Illustration of the photochemical magnetic detection mechanism. A light-induced electron transfer creates an intermediate donor-acceptor pair (D + A) with separate electron spins (dotted arrows). The electron spins move in their local magnetic fields, thereby changing their relative orientation, e.g., from an anti-parallel to a parallel orientation. The relative orientation of the spins, denoted as singlet or triplet spin states, governs what products can be formed and at what rate. Importantly, even a small geomagnetic field (blue) alters the spin motion in a different way on the donor than on the acceptor, in relation to the internal magnetic fields (orange) in both molecules. This results in a change of the singlet to product ratio, as indicated by the blue circles.
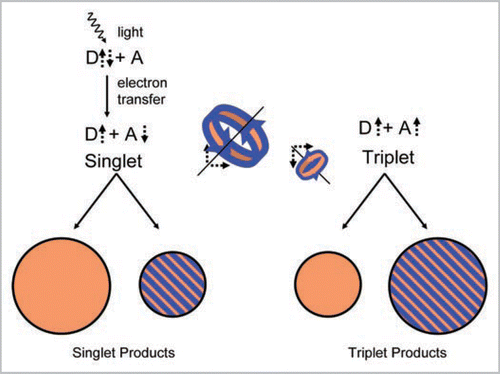
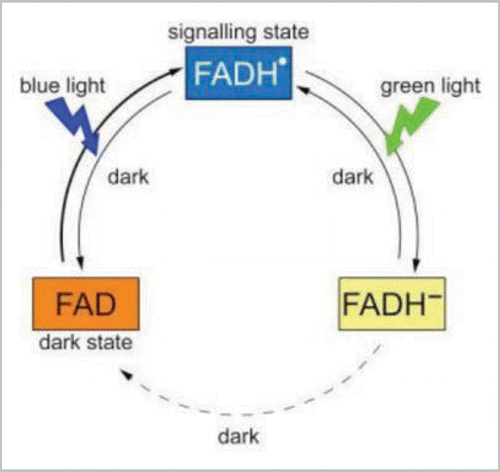
Figure 2 Photocycle of plant cryptochromes. Cryptochromes are bound to a light-absorbing flavin cofactor (FAD) which can exist in three interconvertable redox forms: (FAD, FADHℴ, FADH−). In the dark, cryptochrome is found with flavin in the oxidized state (FAD) which, upon irradiation, undergoes reduction to the FADHℴ form. This reaction requires both electron and proton transfer from nearby amino acid residues in the protein, generating amino acid radicals in turn, which are ultimately reduced through a chain of electron transfer to the protein surface. Further illumination of the flavin radical (by blue or green light) results in formation of the fully reduced FADH− form that is biologically inactive. In the dark, the flavin is spontaneously re-oxidized to restore the fully oxidized resting state through a mechanism that is as yet poorly characterized but involves molecular oxygen and may give rise to superoxide radicals. Therefore both forward and reverse reactions may involve the formation of radical pairs. The degree of cryptochrome activation is governed by the equilibrium reached between the competing forward and reverse reactions under conditions of constant illumination.