Abstract
A tight spatio-temporal coordination of the machineries controlling actin dynamics and membrane remodeling is crucial for a huge variety of cellular processes that shape cells into a multicellular organism. Dynamic membrane remodeling is achieved by a functional relationship between proteins that control plasma membrane curvature, membrane fission and nucleation of new actin filaments. The BAR/ F-BAR-domain-containing proteins are prime candidates to couple plasma membrane curvature and actin dynamics in different morphogenetic processes. Here, we discuss recent findings on the membrane-shaping proteins of the F-BAR domain subfamily and how they regulate morphogenetic processes in vivo.
Figures and Tables
Figure 1 Model for BAR protein function during endocytosis. Schematic drawing, depicting the different steps at which F-BAR proteins are believed to function during endocyosis. (A) Induction of curvature: F-BAR protein dimers bind to and deform the plasma membrane. (B) Invagination/tubulation: By oligomerization, F-BAR proteins form a helical coat around the membrane, thereby stabilizing the invagination. They also might employ actin dynamics to generate the force for membrane invagination. (C) Constriction: (N-)BAR proteins associate with the neck of the tubule, mediating its constriction. (D) Dynamin recruitment: F- and (N-)BAR proteins recruit Dynamin, to also form a coat around the neck of the nascent vesicle, which will ultimately lead to the scission of the vesicle that is also aided by actin polymerization (E) Scission. (F) Vesicle movement: After the scission, WASP and WAVE proteins remain associated with the vesicle via F-BAR proteins, and mediate the formation of an actin tail that propels the vesicle into the cytoplasm.
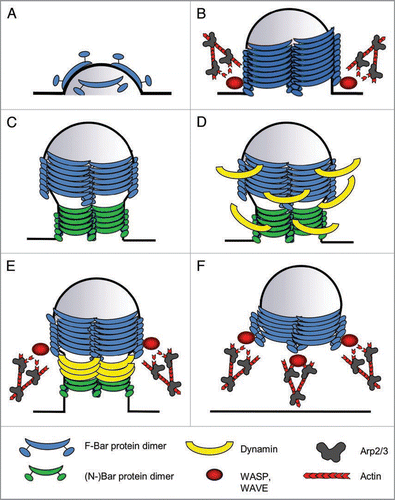
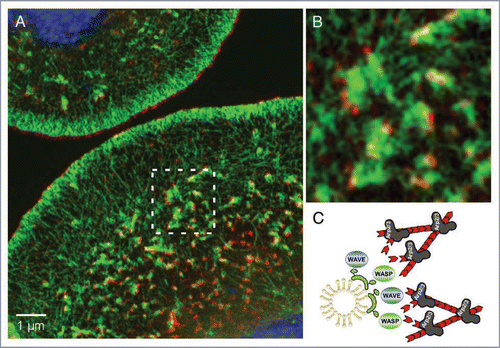
Figure 2 Cip4-marked vesicles localize at the tips of actin tails. (A) Structured Illumination Microscope (SIM) image of Drosophila S2 cells stained for endogenous Cip4 protein (red), F-actin (phalloidin, green). Nuclei were stained with DAPI (blue). (B) A magnified view of (A). Compared to other high-resolution microscopy techniques (e.g., STED) the SIM microscope achieves a resolution approaching 100 nm—double that of a conventional microscope without compromising on dyes or special sample treatment.Citation68