Abstract
Nuclear export of mRNAs is one of the steps critically important for gene expression and different steps of mRNA processing are linked to the export of the mRNA out of the nucleus. This coupling probably provides a quality control mechanism as well as a higher efficiency for the synthesis of mRNAs. The mRNA is synthesized in the nucleus and then exported to the cytoplasm through the nuclear pore complexes (NPCs), which are embedded in the nuclear envelope. The Mex67-Mtr2 complex in yeast and its counterpart Tap-p15 in higher eukaryotes function as an mRNA exporter through the NPC. Some of the DEAD box proteins such as UAP56 and Dbp5 have been implicated in mRNA export also. In this report using the bioinformatics approach we have analyzed the components of the mRNA export machinery in Plasmodium falciparum and also highlighted the salient features of some of the components. Further detailed studies on various components of nuclear mRNA export in Plasmodium falciparum will be essential to understand this important pathway.
The nucleus is a membrane bound compartment and the defining feature of all eukaryotes. It encloses the genome and provides a distinct spatial environment for two of the most fundamental processes of replication and transcription, separating them from the protein translation in the cytoplasm. The access to the nuclear compartment during interphase is restricted due to the presence of a double lipid bilayer called the nuclear envelope (NE). In spite of this physical segregation, there is a constant shuttling of macromolecules between the nucleus and the cytoplasm, so much so that in a growing cell 1 million macromolecules may be transferred between the two compartments every minute.Citation1,Citation2 This directional nucleocytoplasmic exchange is highly specific and regulated, enabling the export of ribosomal RNAs, micro RNA (miRNA), transfer RNA (tRNA), small nuclear RNA (snRNA) and messenger RNA (mRNA) transcripts into the cytoplasm and the import of many proteins required for genome stability, replication, transcription, splicing, export and a host of other processes into the nucleus. This directional transport is a critical cellular process, which influences the gene expression and signal transduction events.
Nuclear export of mRNAs is one of the steps critically important for gene expression. The mRNA is synthesized in the nucleus and then exported to the cytoplasm through the nuclear pore complexes (NPCs), which are large proteinaceous channels embedded in the NE. The overall shape and architecture of the NPC is highly conserved from yeast to higher eukaryotes and these are the only gateway for the transport and exchange of macromolecules between the nucleus and cytoplasm. It is interesting to note that the machinery for export is highly conserved from yeast to humans. NPCs are dynamic octagonally symmetric macromolecular complexes, which mediate nucleocytoplasmic communication and allow shuttling of proteins and ribonucleoprotein complexes (RNPs) and selective transport of cargoes through their central aqueous channel.Citation3,Citation4 NPC are large complexes of the size ranging from 60 Mda in yeast to 125 Mda in vertebrates and harbour multiple copies of 30 to 50 different proteins called Nucleoporins or Nups.Citation5–Citation7 Nups are essential and critical molecular components required for the assembly and function of the NPC. The number of Nups per cell is highly variable and depends on the cell size and activity. Yeast cells have ∼200 NPCs/nucleus and a mature Xenopus oocyte has about 5 × 107 NPCs/nucleus.Citation8 A typical mammalian cell may have as many as 2,000–5,000 NPCs depending on the cell type and growth conditions. To facilitate the import and export of large molecules nuclear transport receptors are required but ions, metabolites and other small molecules of less than 40 kDa can passively diffuse through the aqueous channel of the nuclear pores. Larger macromolecules require soluble nuclear receptors to bind and facilitate the transport of cargo through the NPCs.Citation9,Citation10 Based on the presence or absence of hydrophobic phenylalanine-glycine (FG) repeats, Nups can be categorized into ‘unstructured’ or ‘structured’ forms. The physical barrier imposed by the NPC is due to the presence of a meshwork of FG Nups in the lumen of the channel pores.Citation10,Citation11 FG Nups are natively unfolded subset of Nups, which line the channel pores of the NPC and form a sieve like barrier by interlinking their hydrophobic FG repeat domains.Citation10,Citation12 For mRNA export the binding of export factors to FG motifs (FXFG, GLFG and FG) of NUPs is necessary.Citation13 Nups that are devoid of FG repeats are structured and constitute the backbone of the overall architecture of the NPCs. NPC is made of distinct subcomplexes in yeast and higher eukaryotes. In vertebrates these are the Nup107-160, Nup93 and Nup62 complexes.Citation14 The overall structure and composition of these subcomplexes is conserved during evolution but there is limited sequence conservation between individual Nups of yeast and vertebrates.Citation14
Facilitated nuclear transport is an energy dependent and signal mediated process requiring transport factors belonging to a family of conserved proteins, collectively known as karyopherins (Kaps). These proteins are nuclear transport receptors belonging to the importin β family of HEAT repeat proteins and they are further dependent on the biological activity of a cofactor, the GTPase Ran. Soluble nuclear transport receptors (NTRs) continuously shuttle between the two compartments through the NPCs either alone or together with the cargo macromolecules.Citation15 The transport receptors bind cargo destined for translocation either directly or via an adapter, recognizing certain targeting signals such as Nuclear Localization Signals (NLSs) and Nuclear Export Signals (NESs) to facilitate the transport of cargo through the NPC.Citation16 Moreover, the sequence and the substructural content of NLSs and NESs determine their specificity for various Kaps.Citation17 The passage of the cargo bound receptor complexes through the NPC requires specific interactions with the FG Nups. The FG sequence repeats of Nups serve as a potential docking site for Kaps and these Kaps further provide a biochemical selection mechanism for nucleocytoplasmic transport (NCT).Citation18–Citation21 Moreover, mutations in the transport receptors rendering them defective for binding to the hydrophobic pockets of FG Nups are known to impair their nuclear translocation.Citation22 This NTR—cargo complex is then translocated through the pores of NPC and cargo is released in the destined compartment in an energy dependent manner. Multiple independent import and export pathways have a unifying theme where NTRs perform multiple roles of cargo recognition, translocation through the nuclear pore and cargo release in the destined compartments. Two nucleotide states of the small GTPase Ran, which is predominantly present in the nucleus, play an important role in the directionality of NCT. The binding of RanGTP triggers the association of exportin: cargo complexes and the dissociation of importin: cargo complexes. In the nucleus, RanGTP functions to release the cargo from its import receptor by binding to the receptor itself.Citation23 In the nucleus, exportins bind to their cargo in the presence of GTP and this complex is transported back into the cytoplasm. The GTP hydrolysis at the cytoplasmic NPC face promotes the disassembly of the export receptor-cargo complex in the cytoplasm and this causes the recycling of the export receptor and restores the cytoplasmic RanGDP pool.Citation23
Nuclear microinjection of tRNA in the Xenopus oocyte first revealed that the overall process of RNA export across the nuclear envelope is a carrier mediated translocation process, that is saturable and temperature sensitive.Citation24 Subsequently it became apparent that different RNA species (mRNA, tRNA, miRNA and rRNA) are exported out of the nucleus in a mutually exclusive manner, where one RNA species doesn’t saturate the export of another in a coinjection experiment.Citation25–Citation27 It is well established now that the export of tRNA, miRNA, (sn)RNA and rRNA follows this general scheme that involves exportins of the kaps family and the Ran cycle. But the general mRNA export is mechanistically different as it uses a transport receptor that is unrelated to Kaps and is independent of the Ran cycle. Furthermore numerous additional export factors such as adaptors and release factors cooperate with the mRNA export receptor. The export of tRNA is carried out by the export receptor importin-β family member Exportin-t, which binds tRNA in a RanGTP-dependent manner.Citation28,Citation29 miRNA and rRNA are exported by the karyopherin exportin-5 and Ran/Crm1 export pathway respectively.Citation30–Citation33 The nuclear export of RNA is mediated by proteins but unlike the transport of proteins, members of the Kap superfamily are not involved in mRNA export. mRNA transcripts are transported from the nucleus to the polysomes in the cytoplasm in the form of large ribonucleoprotein (RNP) complexes.Citation34,Citation35 There are constant changes in mRNP composition as these mRNPs progress from synthesis to processing and finally to export.
Plasmodium falciparum is a eukaryotic, unicellular protozoan parasite, which causes the most fatal form of malaria. The disease is a major socio-economic burden on the developing countries causing 300–600 million clinical cases and 2–3 million deaths annually.Citation36,Citation37 With the completion of the genome sequencing in 2002, there has been a surge in the efforts for genome annotation. P. falciparum contains about 5,300 putative protein coding genes, of which only 2,060 have been annotated by sequence similarity and manual curation.Citation38,Citation39 In silico methods for the purpose of gene annotation hold much promise as the classical tools of genetics and biochemistry have been slow to yield results. The A-T richness of the P. falciparum further hampers the gene annotation. Little is known about mRNA export in general and even less is known about mRNA export in P. falciparum. Most of the information about mRNA export has been deciphered using yeast and other higher eukaryotes as the model systems. In this report using the tools of bioinformatics we have identified the components of the mRNA export pathway in P. falciparum. These components were identified mainly by the analysis of the genome using BLAST and profile-Hidden Markov Model searches. Most of the components of the mRNA export pathway are conserved in P. falciparum and other Plasmodium species but there are a few peculiarities which are described in respective sections.
Components of the mRNA Export Machinery in P. falciparum
SR proteins and the TREX complex. The SR (Ser/Arg-rich) proteins are one such family of splicing factors, which can serve as adaptors proteins by recruiting the export factor TAP/NXF to mRNA destined for export.Citation40 These SR proteins contain one or two RNA recognition motifs (RRMs) and Ser/Arg/-rich domain that can be phosphorylated at multiple positions. Ubiquitously found protein complexes like the TREX (transcription/export complex) are involved in coupling activities like the transcript elongation, the splicing and the export.Citation41 TREX complex is a multisubunit complex consisting of THO (suppressor of the transcriptional defect of Hpr1 by overexpression) subcomplex, Tex1 and two additional export factors UAP56 (Sub2 in yeast and UAP56 in metazoa) and Yra1p or ALY/REF adaptor (Yra1 in yeast and ALY or REF in metazoa).Citation42,Citation43 The components of the TREX complex such as Hpr1p are able to recruit the export receptor to the mRNAs by recruiting the export factors Aly and UAP56.Citation43 UAP56 is loaded by components of the TREX complex and it in turn recruits the export receptor TAP/Mex67 by the interaction with ALY/REF.Citation40,Citation44
TREX complex is a highly conserved protein complex involved in the export of mRNA in eukaryotes. In yeast, the TREX complex is made up of a THO subcomplex consisting of Tho2, Hpr1, Mft1, Thp2, and the export factors Sub2 and Yra1 along with Tex1, a protein of unknown function.Citation42 The TREX complex is itself loaded cotranscriptionally to the elongating transcript and is further responsible for recruiting many proteins involved in the process of splicing and mRNA export. SR proteins like Gbp2, Hrb1 and Npl3, are recruited to mRNA by the TREX complex.Citation42 The components of the TREX (THO) complex are responsible for recruiting Grb2 and Hrb1 but not Npl3, which is recruited independently by the interaction with components of the cap binding complex. The shuttling SR proteins apart from carrying out splicing are able to couple splicing and export by acting as export adapters for the mRNA export factors.Citation40 Npl3 has been shown to provide directionality to the process of export by undergoing a cycle of phosphorylation and dephosphorylation events as described previously.
BLAST search (www.ncbi.nlm.nih.gov) using the S. cerevisiae proteins Npl3 and Gbp2 as query in the P. falciparum database “PlasmoDB” (www.plasmodb.org/) revealed two proteins with PlasmoDB number PF10_0217 and PF10_0068 respectively. These proteins PF10_0217 and PF10_0068 are annotated in the PlasmoDB as pre-mRNA splicing factor and RNA binding protein respectively. Moreover, using bioinformatics approaches we were not able to detect the homologue of the SR protein Hrp1 in P. falciparum. PF10_0217 and PF10_0068 contain 538 and 246 amino acids and show considerable similarity to the yeast proteins Npl3 and Gbp2 respectively. The expression data reported in PlasmoDB show that both of these proteins are expressed in all the developmental stages of the parasite. PfNpl3 (PF10_0217) homologue contains two RRMs in the N terminal region of the protein () similar to its human homologue (). PfGbp2 (PF10_0068) homologue surprisingly has only two RRMs (), whereas the corresponding human homologue, heterogeneous nuclear ribonucleoprotein has three RRMs () and is quite larger in size than its counterpart in apicomplexans. The difference in the homologous proteins of P. falciparum and other higher eukaryotes is due to the divergence during the course of evolution. The structural modelling of the RRMs of the PfGbp2 was done using the RNA binding protein Fir as the template.Citation45 The results of this modelling show that there is significant structure conservation in the RRMs of the two proteins as the two structures are completely superimposable ().
In P. falciparum, although some of the components of the TREX complex like Tho2, UAP56 (PfU52) and REF are present, but using the bioinformatics approach we were unable to detect the rest of the components of the TREX complex. It has been reported previously that all the components of the TREX complex are not thoroughly conserved across the various species like homologues of Mft1 and Thp2 have not been found in Drosophila and humans. Moreover, the homologues of yeast Hpr1 and Tex1 are present in Drosophila and humans. Using the bioinformatics approach we were unable to detect the orthologous proteins in P. falciparum. It might be possible that some other proteins are playing the role of TREX homologues in P. falciparum and therefore the TREX complex of this organism is slightly different from the other higher eukaryotes. Using the yeast Tho2 protein sequence as the query, the homologue with the PlasmoDB number PFL2390c (PfTho2) was identified in the P. falciparum genome. PfTho2 is unusually long and contains 2,932 amino acids and shows ∼27% identity and ∼49% homology to the yeast Tho2, which is only 1,597 amino acid long. Tho2 in various species lacks a conserved domain and therefore no conserved domain was observed in PfTho2 using the InterProSan tool (www.ebi.ac.uk). Moreover, PfTho2 is unannotated and has been described as a conserved hypothetical protein of unknown function in PlasmoDB (www.plasmodb.org). The expression data reported in PlasmoDB show that this protein is expressed in all the developmental stages of the parasite. The corresponding homologues in P. vivax (PVX_101385) and P. yoelii (PY01809) are also unannotated hypothetical proteins.
UAP56 is a bona fide splicing factor involved in the export of mRNA transcripts. It directly interacts with the N and C terminal of ALY to recruit it to the spliced mRNPs.Citation46 Apart from Tho2, UAP56 homologue (PfU52) and Ref/Aly (described separately) are also present in the P. falciparum’s TREX complex.Citation47 UAP56 is a member of DEAD box family of RNA helicase, involved in the ATP dependent assembly of spliceosome. UAP56 is an essential protein, that has been implicated in the export of mRNA and it has been shown recently that P. falciparum homologue is an RNA dependent ATPase and it also has a role in the splicing processes.Citation47,Citation48 It was first identified as an interacting partner of the U2AFCitation65 in yeast two hybrid screens for proteins interacting with U2AFCitation65.Citation49 In yeast it was observed that there is a rapid accumulation of poly(A) RNA upon shifting of the temperature sensitive Sub2/UAP56 mutants to non permissive temperatures.Citation50 In Drosophila also double-stranded RNA (dsRNA) mediated depletion of HEL/UAP56 leads to growth inhibition and robust accumulation of poly(A) RNAs in the nucleus.Citation51 The structural modeling of the PfU52 () was done using the human UAP56 () as the template and it was observed that although the overall structure is conserved but insertions in the protein tend to loop out ().Citation52
REF (RNA and Export Factor Binding Proteins) Family
ALY/REF (Yra1p in yeast) is an evolutionary conserved family of hnRNP-like proteins, called REF (RNA and export factor binding proteins), which plays the role of an adaptor protein between REF and mRNPs. Yra1p was originally identified as a yeast nuclear protein which exhibits RNA annealing activity.Citation53 ALY/REF is also a key component of the EJC (exon junction complex) as it remains peripherally associated with the EJC core.Citation54 At least one member of the REF family is present in higher eukaryotes. Multiple members of the REF family have been reported only in Mus musculus, Xenopus laevis, Caenorhabditis elegans, Schizosaccharomyces pombe and Saccharomyces cerevisiae.Citation55 In Saccharomyces cerevisiae the two members of the REF family are known by the names of Yra1 and Yra2. Yra2p when overexpressed is able to complement the deletion of YRA1 in vivo, suggesting that these proteins have redundant functions.Citation43 In P. falciparum PSI-BLAST56 search (www.ncbi.nlm.nih.gov) using the full length protein of yeast Yra1 as a query revealed an ORF with PlasmoDB number PFF0760w, showing ∼28% identity and ∼55% homology with the yeast protein. Interestingly, PFF0760w shows even more similarity to its human homologue ALY, showing ∼34% identity and ∼62% homology. This protein (PFF0760w) has been annotated in PlasmoDB (www.plasmodb.org) as a putative RNA and export factor binding (REF) protein, whereas the corresponding homologues in P. vivax (PVX_113920) and P. knowlesi (PKH_113330) are annotated as RNA binding proteins. The expression data reported in PlasmoDB show that this protein is expressed in all the developmental stages of the parasite. Orthologues of P. falciparum REFp (PlasmoDB number PFF0760w) are annotated as hypothetical proteins in P. yoelii (PY05533) and P. berghei (PB102089.00.0). Although only one member of the REF family is present in the P. falciparum and P. vivax but our bioinformatics analysis reveals that two members of the REF family are present in both P. yoelii (PY07541 and PY05533) and P. berghei (PB102089.00.0 and PB406000.00.0). Surprisingly, the proteins of the REF family from Plasmodium species range between 100–160 amino acids and are smaller than their eukaryotic counterparts, which contain about 200–300 amino acids. Moreover, REF family proteins are characterized by the presence of a central RNA binding domain (RBD), which is flanked on both the sides by two conserved N and C terminal domains.Citation43,Citation55,Citation57 The conserved N and C terminals of the REF proteins are separated by variable insertions of positively charged amino acids.Citation55 In Plasmodium species, the RBD of the REF family is located more towards the C terminal end with a conserved N terminal domain. The C terminal domain of REF in apicomplexans is inconspicuous and is almost fused to the RBD (). The structure of PfREF was modelled and the results show that the structure is almost similar to the RNA binding motif of the template 2E5H (). The structure of the template was downloaded from the MMDB database.Citation58
Tap-p15 Pathway
To transport mRNPs through the NPCs, the yeast Mex67-Mtr2 complex and the homologous metazoan TAP-p15 complex (also known as NXF1-NXT1) function as general mRNA export receptors. Mex67 and TAP bind poly(A) RNA and show genetic and biochemical interactions with the components of the NPC, thus exhibiting characteristic features of the export receptors.Citation59 In yeast, the functionally homologous Mex67 partner in RNA export is Mtr2 whereas in higher eukaryotes Tap and p15 form a heterodimer complex.Citation59 Tap/Mex67 is the principal export factor involved in eukaryotes which mediates the export of the bulk of mRNA from the nucleus.Citation60 In yeast there is a single Tap/NXF homologue known as Mex67 but in higher eukaryotes there are multiple members of the NXF family formed by separate gene duplication events.Citation61 Tap/Mex67 is a modular protein having three distinct domains: a leucine rich repeat domain (LRR), a NTF2 like middle domain and a C terminal UBA (ubiquitin associated fold).Citation62 Mex67p heterodimerizes with Mtr2p with the help of its NTF2 like fold to form heterodimer complex.Citation62 In P. falciparum database PF14_0305 is annotated as the homologue of the nuclear mRNA export factor TAP. This homologue (PF14_0305) shows ∼21% identity and significant similarity (1.4e-05) with the yeast export factor Mex67. The corresponding homologues of TAP in P. vivax (PVX_084925) and P. berghei (PB300366.00.0) are annotated as conserved hypothetical proteins. PF14_0305 and PVX_084925 have a very long N terminal region with insertions of arginine and lysine and are about twice the size of the other eukaryotic counterparts. The modular architecture of PF14_0305 (PfTAP homologue) is not clear as the NTF2 and UBA domains are not apparent in the bioinformatics analysis using the InterProScan sequence search software.Citation63 The absence of the modular architecture in PF14_0305 might be due to the sequence divergence during the course of evolution. It is important to note that the Nxt1/P15 homologue is also absent in P. falciparum and other apicomplexans as revealed by the PSIBLAST search. Nxt1 homologues are apparently present in fungi and other higher eukaryotes but they are absent in protozoans.Citation64
Remodelling ATPase Dbp5 and its Substrates
It has been suggested that two DEAD-box helicases Dbp5 and UAP56 play key roles in the process of RNA export and are indispensable for this export.Citation65–Citation67 RNA helicases are ubiquitously involved in each step of RNA metabolism. Dbp5 determines the overall directionality of the process of mRNA export. The activation of Dbp5 in turn is controlled by IP6 (Inositol hexaphosphate)—bound Gle1 at the cytoplasmic side of the NPC.Citation68,Citation69 This activation most likely facilitates the remodeling of mRNP protein composition during directional transport and provides energy for transport cycles.Citation68,Citation69 The weak ATPase activity of Dbp5 is stimulated by the cellular cofactors like Gle1 and IP6.Citation68,Citation69 Recombinant CTD (C terminal domain) of Gle1 activates Dbp5 by decreasing the Km of the enzyme for ATP approximately about six fold and side by side increases the Kcat about four-fold, resulting in the overall increase in the kinetic efficiency of the enzyme.Citation69 The stimulation of Dbp5 is in turn required for RNA unwinding and displacement of proteins from RNA-protein complexes. Gle1 binds to Nup42 and anchors to the cytoplasmic fibrils of NPC at a site, which is juxtaposed to the Dbp5-Nup159 binding site.Citation70,Citation71
Dbp5 is a yeast RNA helicase involved in remodelling RNPs extruding out of the NPC by removing Mex67 and other hnRNP like proteins from the exported mRNP.Citation67,Citation72 In a Dbp5 mutant, there is an increased association of Mex67 with the mRNAs, suggesting a direct role of this protein in RNP remodelling at the nuclear rim.Citation67 Most of the physiological substrates of Dbp5, like Mex67 and other mRNP bound proteins tend to accompany the mRNP into the cytoplasm through the nucleoplasm but are not found in association with the polysomes.Citation73 All of these results suggest that these accompanying proteins are removed upon entry into the cytoplasm from the NPC, just prior to translation. It has been shown that the bound proteins are displaced by the ADP bound form of Dbp5.Citation72 Recently, it has been shown that Nab2p is one of the physiological substrate for Dbp5p. Moreover, Nab2p bound RNPs are targeted by Dbp5p at the cytoplasmic face of the nuclear pore, where the RNP bound proteins like Nab2 are removed from the mRNP by Dbp5p.Citation72
In P. falciparum genome the gene with PlasmoDB number PF14_0563 is the homologue of Dbp5/Rat8 and is annotated in PlasmoDB (www.plasmodb.org) as a putative “DEAD-box RNA helicase”.Citation74,Citation75 Orthologues of Dbp5p are also annotated as “DEAD-box RNA helicase” in other species like P. vivax (PVX_117510), P. yoelii (PY06529), P. chaubadi (PC000054.02.0) and P. berghie (PB000472.01.0). BLAST search using the yeast Dbp5 as the query, revealed the presence of proteins which are very closely related in other apicomplexan species like Cryptosporidium parvum (XP_627359), Babesia bovis (XP_001609208), Toxoplasma gondii (EEB02796), Theileria parva (XP_765582), Theileria annulata (XP_954649) and Cryptosporidium hominis (XP_666618).Citation56 We are characterizing the Dbp5 homologue from P. falciparum and the results will be published separately (Mehta and Tuteja, manuscript in preparation).
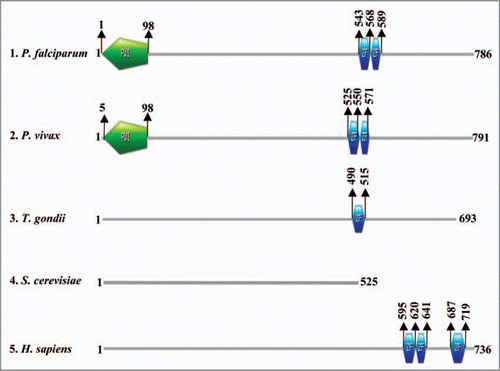
Using the protein sequences of Nab2 (yeast) and the human homologue (Zinc finger CCCH domain-containing protein 14/NY-REN-37) as query a Nab2 homologue was detected in PlasmoDB (www.plasmodb.org). The gene with PlasmoDB number PFF1110c is P. falciparum Nab2 homologue and it shows ∼46% homology with the yeast protein and ∼63% homology (4.5e-17) with the human homologue NY-REN-37. It is important to note that this gene (PFF1110c) is unannotated and has been described as conserved hypothetical protein in the database. The orthologous proteins of P. vivax (PVX_114270), P. yoelii (PY03499) and P. berghie (PB000664.00.0) are also described as hypothetical proteins. Furthermore the results of the InterProScan (www.ebi.ac.uk) of the corresponding (PFF1110c, PfNab2) protein revealed the presence of two Zinc finger CCCH type motif (from amino acid 543 to 568 and 569 to 589) and a conserved panther domain from amino acid 525 to 785 (PTHR14738) (). The results of our analysis show that the Zinc finger CCCH type motif is only present in the orthologous proteins of P. vivax, H. sapiens, C. elegans and absent in the S. cerevisiae homologue Nab2. Moreover P. falciparum and P. vivax homologues of Nab2 also contain a PWI domain at their N terminal (). A PWI domain is a novel nucleic acid binding motif and the proteins bearing this motif are involved in the processes like transcription, 5′-end capping, splicing, 3′-end processing (cleavage and polyadenylation), surveillance, turnover, transport and translation.Citation76 Since the PWI domain is absent in the human as well as the yeast orthologues, we propose here that PfNab2 might be playing roles other than the ones played by Nab2 protein of yeast and it might be a key protein involved in the RNA metabolism.
Concluding Remarks
It is well established that the export of mRNA from the nucleus to the cytoplasm takes place through a well defined pathway. Export competent mRNP assembly is coupled to transcription and this mRNP is composed of mRNA and a number of other RNA export factors. These export factors include Mex67-Mtr2, Npl3, poly(A) binding protein (PABP) and the DEAD-box protein Dbp5. Dbp5 is responsible for the release of mRNA from mRNP in the cytoplasm. The knowledge of mRNA export mechanisms in malaria parasites is very limited and in its infancy. Our in silico studies reported here suggest that some variations in RNA export pathway exist in the malaria parasite. We have already initiated the characterization of some of these components such as PABPCitation77,Citation78 and Dbp5 (Mehta and Tuteja, manuscript in preparation) from P. falciparum. The comparative studies reported here have laid the foundation for future work and further detailed studies on the various components of the RNA export machinery in the malaria parasite are essential to better understand this important process.
Figures and Tables
Figure 1 Schematic diagram showing the domain organization of Npl3 homologues (A and B) and Gbp2 homologues (C and D) of Plasmodium falciparum and Homo sapiens. The protein names and species are written on the left hand side in each panel. The domain analysis was done using Scan Prosite at (expasy.org). (E) Superimposed image of the computer based structure model of the RR M domain of Plasmodium falciparum Gbp2 (shown in pink) on the RR M domain of Fir (2qfj) as a parent template (shown in yellow).
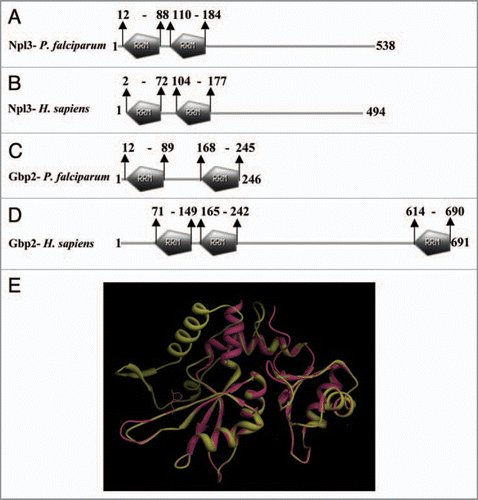
Figure 2 The computer based structure model of (A) PfU52 of Plasmodium falciparum and (B) Human UAP56 (1XTI). (C) The superimposed image.
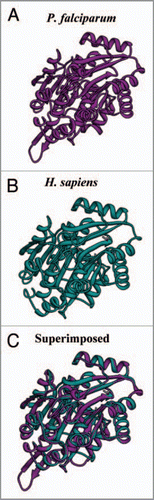
Figure 3 (A) Comparison of the domain organization in the Ref proteins of Higher eukaryotes (i) and apicomplexans (ii). The conserved N and C terminal domains flanking the RR M motif are shown as RE F-N and RE F-C respectively. N-v and C-v represents the N and C terminal variable regions. (The figure has been prepared with the help of information given in ref. 55). (B) Schematic diagram showing the domain organization in the RE F family of (1) Plasmodium falciparum, (2) Plasmodium vivax, (3) Toxoplasma gondii, (4) Sacchromyces cerevisiae and (5) Homo sapiens. The domain analysis was done using Scan Prosite at (http://expasy.org). The protein names and species are written on the left hand side.
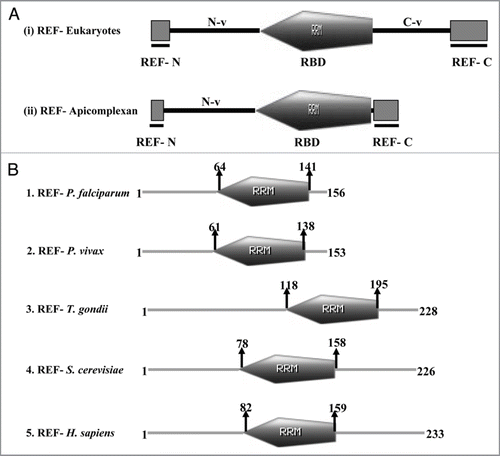
Figure 4 The computer based structure modeling of (A) PfRE F of Plasmodium falciparum based on the (B) 2E5 h-template (C) The superimposed image.
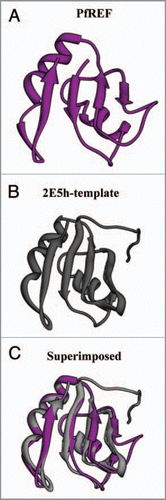
Figure 5 Schematic diagram showing the domain organization in various homologues of Nab2 in (1) Plasmodium falciparum, (2) Plasmodium vivax, (3) Toxoplasma gondii, (4) Sacchromyces cerevisiae and (5) Homo sapiens. The domain analysis was done using Scan Prosite at (http://expasy.org). The names of species are written on the left hand side.
Acknowledgements
The work in RT’s laboratory is supported by grants from Department of Biotechnology, Defence Research and Development Organization and Department of Science and Technology. Infrastructural support from the Department of Biotechnology, Government of India is gratefully acknowledged.
References
- Ribbeck K, Gorlich D. Kinetic analysis of translocation through nuclear pore complexes. EMBO J 2001; 20:1320 - 1330
- Yang W, Gelles J, Musser SM. Imaging of single-molecule translocation through nuclear pore complexes. Proc Natl Acad Sci USA 2004; 101:12887 - 12892
- Davis LI. The nuclear pore complex. Annu Rev Biochem 1995; 64:865 - 896
- Fahrenkrog B, Aebi U. The nuclear pore complex: nucleocytoplasmic transport and beyond. Nat Rev Mol Cell Biol 2003; 4:757 - 766
- Rout MP, Blobel G. Isolation of the yeast nuclear pore complex. J Cell Biol 1993; 123:771 - 783
- Rout MP, Aitchison JD, Suprapto A, Hjertaas K, Zhao Y, Chait BT. The yeast nuclear pore complex: composition, architecture and transport mechanism. J Cell Biol 2000; 148:635 - 651
- Cronshaw JM, Krutchinsky AN, Zhang W, Chait BT, Matunis MJ. Proteomic analysis of the mammalian nuclear pore complex. J Cell Biol 2002; 158:915 - 927
- Gorlich D, Kutay U. Transport between the cell nucleus and the cytoplasm. Annu Rev Cell Dev Biol 1999; 15:607 - 660
- Frey S, Görlich D. A saturated FG-repeat hydrogel can reproduce the permeability properties of nuclear pore complexes. Cell 2007; 130:405 - 407
- Tran EJ, Wente SR. Dynamic nuclear pore complexes: life on the edge. Cell 2006; 125:1041 - 1053
- Frey S, Richter RP, Gorlich D. FG-rich repeats of nuclear pore proteins form a three-dimensional meshwork with hydrogel-like properties. Science 2006; 314:815 - 817
- Denning DP, Patel SS, Uversky V, Fink AL, Rexach M. Disorder in the nuclear pore complex: The FG repeat regions of nucleoporins are natively unfolded. Proc Natl Acad Sci USA 2003; 100:2450 - 2455
- Strasser K, Bassler J, Hurt E. Binding of the Mex67p/Mtr2p heterodimer to FXFG, GLFG and FG repeat nucleoporins is essential for nuclear mRNA export. J Cell Biol 2000; 150:695 - 706
- Schwartz TU. Modularity within the architecture of the nuclear pore complex. Curr Opin Struct Biol 2005; 15:221 - 226
- Fried H, Kutay U. Nucleocytoplasmic transport: taking an inventory. Cell Mol Life Sci 2003; 60:1659 - 1688
- Englmeier L, Fornerod M, Bischoff FR, Petosa C, Mattaj IW, Kutay U. Ran BP3 influences interaction between CRM1 and its export substrates. EMBO Rep 2001; 2:926 - 932
- Friedrich B, Quensel C, Sommer T, Hartmann E, Köhler M. Nuclear localization signal and protein context both mediate importin alpha specificity of nuclear import substrates. Mol Cell Bio 2006; 26:8697 - 8709
- Bayliss R, Leung SW, Baker RP, Quimby BB, Corbett AH, Stewart M. Structural basis for the interaction between NTF2 and nucleoporin FxFG repeats. EMBO J 2002; 21:2843 - 2853
- Bednenko J, Cingolani G, Gerace L. Importin beta contains a COOH-terminal nucleoporin binding region important for nuclear transport. J Cell Biol 2003; 162:391 - 401
- Pemberton LF, Paschal BM. Mechanisms of receptormediated nuclear import and nuclear export. Traffic 2005; 6:187 - 198
- Cushman I, Palzkill T, Moore MS. Using peptide arrays to define nuclear carrier binding sites on nucleoporins. Methods 2006; 39:329 - 341
- Bayliss R, Littlewood T, Strawn LA, Wente SR, Stewart M. GLFG and FxFG nucleoporins bind to overlapping sites on importin-beta. J Biol Chem 2002; 277:50597 - 50606
- Gorlich D, Pante N, Kutay U, Aebi U, BischoV FR. Identification of different roles for RanGDP and RanGTP in nuclear protein import. EMBO J 1996; 15:5584 - 5594
- Zasloff M. tRNA transport from the nucleus in a eukaryotic cell: Carrier-mediated translocation process. Proc Natl Acad Sci USA 1983; 80:6436 - 6440
- Bataille N, Helser T, Fried HM. Cytoplasmic transport of ribosomal subunits microinjected into the Xenopus laevis oocyte nucleus: A generalized, facilitated process. J Cell Biol 1990; 111:1571 - 1582
- Jarmolowski A, Boelens WC, Izaurralde E, Mattaj IW. Nuclear export of different classes of RNA is mediated by specific factors. J Cell Biol 1994; 124:627 - 635
- Pokrywka NJ, Goldfarb DS. Nuclear export pathways of tRNA and 40 S ribosomes include both common and specific intermediates. J Biol Chem 1995; 270:3619 - 3624
- Arts GJ, Fornerod M, Mattaj IW. Identification of a nuclear export receptor for tRNA. Curr Biol 1998; 8:305 - 314
- Kutay U, Lipowsky G, Izaurralde E, Bischoff FR, Schwarzmaier P, Hartmann E, et al. Identification of a tRNA-specific nuclear export receptor. Mol Cell 1998; 1:359 - 369
- Hurt E, Hannus S, Schmelzl B, Lau D, Tollervey D, Simos G. A novel in vivo assay reveals inhibition of ribosomal nuclear export in Ran-cycle and nucleoporin mutants. J Cell Biol 1999; 144:389 - 401
- Ho JHN, Kallstrom G, Johnson AW. Nmd3p is a Crm1p-dependent adapter protein for nuclear export of the large ribosomal subunit. J Cell Biol 2000; 151:1057 - 1066
- Yi R, Qin Y, Macara IG, Cullen BR. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev 2003; 17:3011 - 3016
- Lund E, Guttinger S, Calado A, Dahlberg JE, Kutay U. Nuclear export of microRNA precursors. Science 2004; 303:95 - 98
- Dreyfuss G, Matunis MJ, Pinol-Roma S, Burd CG. hnRNP proteins and the biogenesis of mRNA. Annu Rev Biochem 1993; 62:289 - 321
- Daneholt B. A look at messenger RNP moving through the nuclear pore. Cell 1997; 88:585 - 588
- Snow RW, Guerra CA, Noor AM, Myint HY, Hay SI. The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature 2005; 434:214 - 217
- Tuteja R. Malaria-An overview. FEBS J 2007; 274:4670 - 4679
- Gardner MJ, Hall N, Fung E, White O, Berriman M, Hyman RW, et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 2002; 419:498 - 511
- Ginsburg H. Caveat emptor: limitations of the automated reconstruction of metabolic pathways in Plasmodium. Trends Parasitol 2009; 25:37 - 43
- Huang Y, Gattoni R, Stevenin J, Steitz JA. SR splicing factors serve as adapter proteins for TAP-dependent mRNA export. Mol Cell 2003; 11:837 - 843
- Strässer K, Masuda S, Mason P, Pfannstiel J, Oppizzi M, Rodriguez-Navarro S, et al. TREX is a conserved complex coupling transcription with messenger RNA export. Nature 2002; 417:304 - 308
- Chávez S, Beilharz T, Rondón AG, Erdjument-Bromage H, Tempst P, Svejstrup JQ, et al. A protein complex containing Tho2, Hpr1, Mft1 and a novel protein, Thp2, connects transcription elongation with mitotic recombination in Saccharomyces cerevisiae. EMBO J 2000; 19:5824 - 5834
- Zenklusen D, Vinciguerra P, Wyss JC, Stutz F. Stable mRNP formation and export require cotranscriptional recruitment of the mRNA export factors Yra1p and Sub2p by Hpr1p. Mol Cell Bio 2002; 22:8241 - 8253
- Zhou Z, Luo MJ, Straesser K, Katahira J, Hurt E, Reed R. The protein Aly links pre-messenger-RNA splicing to nuclear export in metazoans. Nature 2000; 407:401 - 405
- Crichlow GV, Zhou H, Hsiao HH, Frederick KB, Debrosse M, Yang Y, et al. Dimerization of FIR upon FUSE DNA binding suggests a mechanism of c-myc inhibition. EMBO J 2008; 27:277 - 289
- Sträßer K, Hurt EC. Splicing factor Sub2p is required for nuclear export through its interaction with Yra1p. Nature 2001; 413:648 - 652
- Shankar J, Pradhan A, Tuteja R. Isolation and characterization of Plasmodium falciparum UAP56 homologue: evidence for the coupling of RNA binding and splicing activity by site-directed mutations. Arch Biochem Biophy 2008; 478:143 - 153
- MacMorris M, Brocker C, Blumenthal T. UAP56 levels affect viability and mRNA export in Caenorhabditis elegans. RNA 2003; 9:847 - 857
- Fleckner J, Zhang M, Valcarcel J, Green MR. U2AF65 recruits a novel human DEAD box protein required for the U2 snRNP-branchpoint interaction. Genes Dev 1997; 11:1864 - 1872
- Jensen TH, Patricio K, McCarthy T, Rosbash M. A block to mRNA nuclear export in S. cerevisiae leads to hyperadenylation of transcripts that accumulate at the site of transcription. Mol Cell 2001; 7:887 - 898
- Gatfield D, Le Hir H, Schmitt C, Braun IC, Köcher T, Wilm M, et al. The DExH/D box protein HEL/UAP56 is essential for mRNA nuclear export in Drosophila. Curr Biol 2001; 11:1716 - 1721
- Cordin O, Minder CM, Linder P, Xu RM. Crystal structure of the human ATP-dependent splicing and export factor UAP56. Proc Natl Acad Sci USA 2004; 101:17628 - 17633
- Portman DS, O’Connor JP, Dreyfuss G. Yra1, an essential Saccharomyces cerevisiae gene, encodes a novel nuclear protein with RNA annealing activity. RNA 1997; 3:527 - 537
- Bono F, Ebert J, Lorentzen E, Conti E. The crystal structure of the exon junction complex reveals how it maintains a stable grip on mRNA. Cell 2006; 126:713 - 725
- Stutz F, Bachi A, Doerks T, Braun IC, Seraphin B, Wilm M, et al. REF, an evolutionary conserved family of hnRNP-like proteins, interacts with TAP/Mex67p and participates in mRNA nuclear export. RNA 2000; 6:638 - 650
- Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, et al. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997; 25:3389 - 3402
- Rodrigues JP, Rode M, Gatfield D, Blencowe BJ, Carmo-Fonseca M, Izaurralde E. REF proteins mediate the export of spliced and unspliced mRNAs from the nucleus. Proc Natl Acad Sci USA 2001; 98:1030 - 1035
- Wang Y, Addess KJ, Chen J, Geer LY, He J, He S, et al. MMDB: annotating protein sequences with Entrez’s 3D-structure database. Nucleic Acids Res 2007; 35:298 - 300
- Segref A, Sharma K, Doye V, Hellwig A, Huber J, Luhrmann R, et al. Mex67p, a novel factor for nuclear mRNA export, binds to both poly(A)+ RNA and nuclear pores. EMBO J 1997; 16:3256 - 3271
- Hurt E, Strässer K, Segref A, Bailer S, Schlaich N, Presutti C, et al. Mex67p mediates the nuclear export of a variety of pol II transcripts. J Biol Chem 2000; 275:8361 - 8368
- Herold A, Suyama M, Rodrigues JP, Braun IC, Kutay U, Carmo-Fonseca M, et al. TAP/NXF1 belongs to a multigene family of putative RNA export factors with a conserved modular structure. Mol Cell Biol 2000; 20:8996 - 9008
- Suyama M, Doerks T, Braun IC, Sattler M, Izaurralde E, Bork P. Prediction of structural domains of TAP reveals details of its interaction with p15 and nucleoporins. EMBO Rep 2000; 1:53 - 58
- Hunter S, Apweiler R, Attwood TK, Bairoch A, Bateman A, Binns D, et al. InterPro: the integrative protein signature database. Nucleic Acids Res 2009; 37:211 - 215
- Mans BJ, Anantharaman V, Aravind L, Koonin EV. Comparative genomics, evolution and origins of the nuclear envelope and nuclear pore complex. Cell Cycle 2004; 3:1612 - 1637
- Tseng SS, Weaver PL, Liu Y, Hitomi M, Tartakoff AM, Chang TH. Dbp5p, a cytosolic RNA helicase, is required for poly (A)+ RNA export. EMBO J 1998; 17:2651 - 2662
- Zhao J, Jin SB, Bjorkroth B, Wieslander L, Daneholt B. The mRNA export factor Dbp5 is associated with Balbiani ring mRNP from gene to cytoplasm. EMBO J 2002; 21:1177 - 1187
- Lund MK, Guthrie C. The DEAD-box protein Dbp5p is required to dissociate Mex67p from exported mRNPs at the nuclear rim. Mol Cell 2005; 20:645 - 651
- Alcazar-Roman AR, Tran EJ, Guo S, Wente SR. Inositol hexakisphosphate and Gle1 activate the DEAD-box protein Dbp5 for nuclear mRNA export. Nature Cell Biol 2006; 8:711 - 716
- Weirich CS, Erzberger JP, Berger JM, Weis K. The N-terminal domain of Nup159 forms a β-propeller that functions in mRNA export by tethering the helicase Dbp5 to the nuclear pore. Mol Cell 2004; 16:749 - 760
- Rayala HJ, Kendirgi F, Barry DM, Majerus PW, Wente SR. The mRNA export factor human Gle1 interacts with the nuclear pore complex protein Nup155. Mol Cell Proteomics 2004; 3:145 - 155
- Kendirgi F, Barry DM, Griffis ER, Powers MA, Wente SR. An essential role for hGle1 nucleocytoplasmic shuttling in mRNA export. J Cell Biol 2003; 160:1029 - 1040
- Tran EJ, Zhou Y, Corbett AH, Wente SR. The DEAD-box protein Dbp5 controls mRNA export by triggering specific RNA:protein remodeling events. Mol Cell 2007; 28:850 - 859
- Windgassen M, Sturm D, Cajigas IJ, Gonzalez CI, Seedorf M, Bastians H, et al. Yeast shuttling SR proteins Npl3p, Gbp2p and Hrb1p are part of the translating mRNPs, and Npl3p can function as a translational repressor. Mol Cell Biol 2004; 24:10479 - 10491
- Tuteja R, Pradhan A. Unraveling the ‘DEAD-box’ helicases of Plasmodium falciparum. Gene 2006; 376:1 - 12
- Tuteja R. Genome wide identification of Plasmodium falciparum helicases: a comparison with human host. Cell Cycle 2010; 9:104 - 121
- Szymczyna BR, Bowman J, McCracken S, Pineda-Lucena A, Lu Y, Cox B, et al. Structure and function of the PWI motif: a novel nucleic acid-binding domain that facilitates pre-mRNA processing. Genes Dev 2003; 17:461 - 475
- Tuteja R. Identification and bioinformatics characterization of translation initiation complex eIF4F components and poly(A)-binding protein from Plasmodium falciparum. Communicative & Integrative Biology 2009; 2:1 - 16
- Tuteja R, Pradhan A. Isolation and functional characterization of eIF4F components and poly(A)-binding protein from Plasmodium falciparum. Parasitology International 2009; 58:481 - 485