
Abstract
The role of photosynthesis in plant defense is a fundamental question awaiting further molecular and physiological elucidation. To this end we investigated host responses to infection with the bacterial pathogen Xanthomonas axonopodis pv. citri, the pathogen responsible for citrus canker. This pathogen encodes a plant-like natriuretic peptide (XacPNP) that is expressed specifically during the infection process and prevents deterioration of the physiological condition of the infected tissue. Proteomic assays of citrus leaves infected with a XacPNP deletion mutant (∆XacPNP) resulted in a major reduction in photosynthetic proteins such as Rubisco, Rubisco activase and ATP synthase as a compared with infection with wild type bacteria. In contrast, infiltration of citrus leaves with recombinant XacPNP caused an increase in these host proteins and a concomitant increase in photosynthetic efficiency as measured by chlorophyll fluorescence assays. Reversion of the reduction in photosynthetic efficiency in citrus leaves infected with ∆XacPNP was achieved by the application of XacPNP or Citrus sinensis PNP lending support to a case of molecular mimicry. Finally, given that ∆XacPNP infection is less successful than infection with the wild type, it appears that reducing photosynthesis is an effective plant defense mechanism against biotrophic pathogens.
Figures and Tables
Figure 1 Changes in photosynthetic proteins during bacterial infections and XacPNP treatment. Protein spots from 2-DE SDS-PAGE of proteins from citrus leaves stained with Coomassie blue. Citrus leaves were infiltrated with XacWT, ΔXacPNP (107 CFU/ml) and 5 µM XacPNP pure protein (+XacPNP). After 3 days of bacterial infections or 30 minutes after infiltration with recombinant protein, total plant proteins were extracted and subjected to the proteomics analysis. As control, citrus leaves were infiltrated with Tris 50 mM.

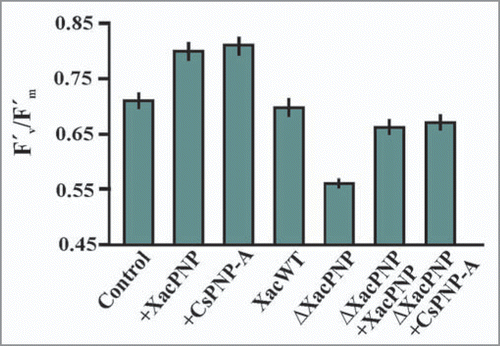
Figure 2 Effect of PNPs in effective quantum efficiency of photosystem II in host leaves. Chlorophyll fluorescence was measured by an 0.8 s saturating pulse at 5,000 mmol m−2 s−1 in leaves infiltrated with 5 µM PNPs, Xac wild type, ΔXacPNP and ΔXacPNP (107 CFU/ml) complemented with XacPNP and CsPNP-A. In the control, citrus leaves were infiltrated with Tris 50 mM. The results are the mean of three replicates and error bars represent the standard deviations.
Addendum to: and