
Abstract
The question of whether or not humans possess "free will" to control their own actions has preoccupied generations of scholars in the realm of Religion and Science. In Jewish theological literature, for instance, the contradiction between God's unlimited power of our destiny and Man's own "free will" represents one of the most challenging paradoxes of religious thought that is beyond our understanding. In Jewish philosophy, "Everything is foreseen; yet free will is given" (Rabbi Akiva, Pirkei Avoth 3:15), and this is where the paradox lies. But if we assume that humans do possess "free will" to make their own spontaneous choices, then this ability must be embedded in our brains. A crucial question then comes to mind: is "free will" unique to humans or is it a trait shared by other organisms as well?
Figures and Tables
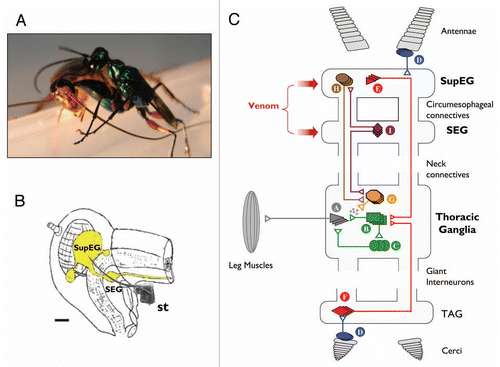
Figure 1 (A) The parasitoid Jewel Wasp A. compressa stings its cockroach prey inside the head. (B) Schematic drawing of the cerebral nervous system (yellow) inside the cockroach's head capsule. the wasp's stinger (st., scanning electron micrograph drawn to scale) reaches to inject venom into both cerebral ganglia, namely the supra-esophageal ganglion (SupEG) and sub-esophageal ganglion (SEG). Scale bar: 0.5 mm. (C) Current model of the neurophysiological events leading to venom-induced hypokinesia in cockroaches stung by the Jewel Wasp. Schematic and simplified drawing of a cockroach nervous system, depicting circuitries that affect walking-related behaviors. The walking pattern generator that orchestrates leg movements is located in the thorax. It consists of motor neurons (A) innervating leg muscles, afferent neurons from sensory structures on the legs (not shown) and type-A thoracic interneurons (TIAs; B), which synapse onto the motor neurons directly and indirectly via local interneurons (C). The TIAs receive inputs from several interneurons. For example, sensory neurons (D) in the antennae or cerci recruit descending (E) or ascending (F) Giant Interneurons (GIs) in the SupEG and terminal abdominal ganglion (TAG), respectively. The GIs converge directly onto the TIAs to evoke escape responses. In addition, neurons of the pattern generator receive input from thoracic neuromodulatory cells (G). One example of these is the thoracic dorsal unpaired median (DUM) neurons, which secrete octopamine and modulate the efficacy of premotor-to-motor synapses. The neuromodulatory cells, in turn, receive tonic regulatory input from interneurons descending from the supra-esophageal ganglion (SupEG) (H) and sub-esophageal ganglion (SEG) (I). This tonic input affects the probability of occurrence of specific motor behaviors by modulating the different thoracic pattern generators directly (not shown) or indirectly (H and I). The wasp, A. compressa, injects its venom cocktail directly into both cerebral ganglia to modulate specific, yet unidentified cerebral circuitries. The current hypothesis states that in the SEG, the venom suppresses the activity of neuromodulatory neurons (I), presumably SEG-DUM neurons, which (1) ascend to the SupEG to regulate descending neuromodulatory (probably octopaminergic) neurons (H), and/or (2) descend to the thorax to regulate locomotory CPGs directly (not shown) or indirectly through thoracic neuromodulatory (probably DUM) neurons (G). Hence, the venom injected into the cerebral ganglia decreases the overall excitatory input to the thoracic walking pattern generator. As a result, walking-related behaviors are specifically inhibited and stimuli to the antennae or cerci fail to evoke normal escape responses. Figure modified from Libersat et al. (2009).Citation21
Addendum to: