Abstract
In humans, there is an interdependent relationship between aging and immune system function, with each process affecting the outcome of the other. Aging can trigger immune system dysfunction, and alterations in the immune response can in turn affect human lifespan. Genetic experiments in model organisms such as C. elegans and Drosophila have led to the identification of numerous genes and signaling pathways that can modulate organismal lifespan and immune system function. Importantly, many of these signaling pathways exhibit conserved function in multiple species, including mammals, suggesting that the research in these simpler models could one day pave the way for the modulation of aging and immunity in humans. Here, we review the recent progress in our understanding of aging, innate immunity, and the interaction between these two processes using these simple model systems. Additionally, we discuss what this may tell us about aging and the innate immune system in humans.
Acknowledgements
I thank Danielle Garsin, Queelim Ch'ng and Jennifer Kemp for critical reading of this manuscript.
Figures and Tables
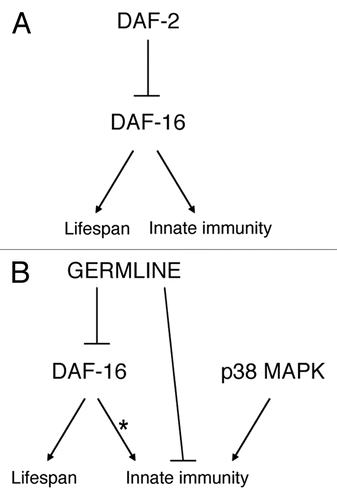
Figure 1 Signaling pathways that regulate longevity in C. elegans. The daf-2 insulin/IGF-1-like signaling pathway (A) and germline proliferation (B) both regulate the DAF-16/FOXO family transcription factor, which in turn regulates nematode longevity.
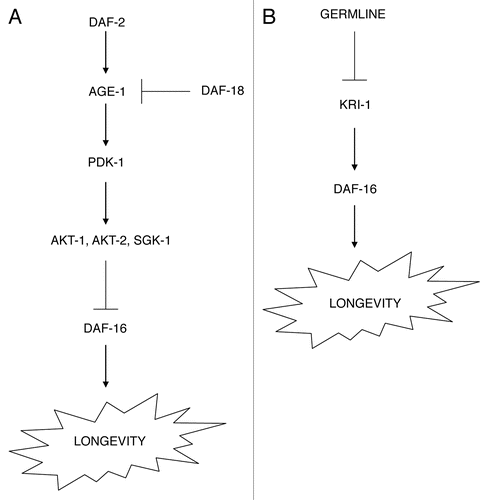
Figure 2 Innate immunity regulation in C. elegans. Three key signaling pathways regulate innate immunity in C. elegans. The p38 MAPK cascade functions in the intestine and epidermis to regulate antimicrobial gene expression, and includes TIR-1, a protein with a conserved TIR domain, the MAPKKK NSY-1, the MAPKK SEK-1 and the MAPK PMK-1 (A). How this pathway senses pathogens is still unclear, but the specificity of the nematode innate immune response suggests that several pathogen recognition receptors (PRRs) exist in the genome. The TGFβ family member DBL-1 is synthesized in neurons and controls expression of antimicrobial genes in the epidermis (B). DBL-1 signals via the TGFβ receptor (composed of DAF-4 and SMA-6) and a downstream SMAD (SMA-3). Neuronal expression of the insulin-like peptide INS-7 also regulates antimicrobial gene expression non-autonomously, acting on the DAF-2 receptor and downstream signaling components, including DAF-16 in the intestine (C).
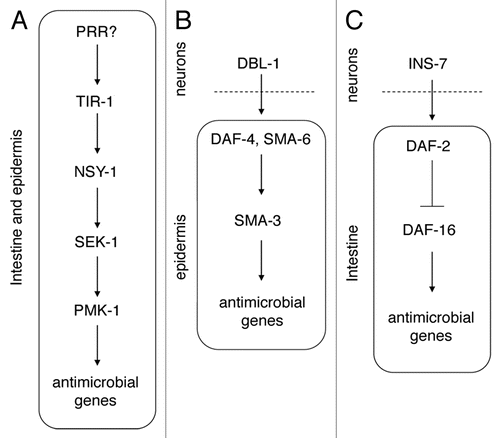
Figure 3 Model for the regulation of lifespan and innate immunity. The daf-2 (A) and germline (B) pathways both regulate lifespan and innate immunity in C. elegans. The effect of daf-2 on both lifespan and innate immunity depends on the DAF-16 transcription factor (A). Similarly, the effect of the germline on lifespan also requires daf-16. However, depending on the growth conditions of the pathogen and the nematodes, the effect of the germline on innate immunity may or may not depend on DAF-16 (B). The germline also acts in parallel to a p38 MAPK pathway to regulate innate immunity. This figure was originally published in Alper et al. “The Caenorhabditis elegans germ line regulates distinct signaling pathways to control lifespan and innate immunity.” J Biol Chem 2010; 285:1822–8.