Abstract
The integrin-mediated adhesion machinery is the primary cell-matrix adhesion mechanism in Metazoa. The integrin adhesion complex, which modulates important aspects of the cell physiology, is composed of integrins (α and β subunits) and several scaffolding and signaling proteins. Integrins appeared to be absent in all non-metazoan eukaryotes so far analyzed, including fungi, plants, and choanoflagellates, the sister-group to Metazoa. Thus, integrins and, therefore, the integrin-mediated adhesion and signaling mechanism was considered a metazoan innovation. Recently, a broad comparative genomic analysis including new genome data from several unicellular organisms closely related to fungi and metazoans shattered previous views. The integrin adhesion and signaling complex is not specific to Metazoa, but rather it is present in apusozoans and holozoan protists. Thus, this important signaling and adhesion system predated the origin of Fungi and Metazoa, and was subsequently lost in fungi and choanoflagellates. This finding suggests that cooption played a more important role in the origin of Meta-zoa than previously believed. Here, we hypothesize that the integrin adhesome was ancestrally involved in signaling.
Both cell adhesion and cell signaling, which are often correlated, are essential mechanisms for metazoan multicellularity.Citation1 Thus, elucidating the origin and evolution of those processes is key to further our understanding of metazoan origins. It has been inferred that the most ancestral cell junctions in metazoans are spot adherens junctions and focal adhesions.Citation2 The former are important for cell-cell adhesion and are molecularly based on cadherins, coupled with the actin cytoskeleton through catenins. On the other hand, focal adhesions are essential for cell-extracellular matrix (ECM) connection and are molecularly based on the integrin adhesion complex also linked to the actin cytoskeleton.
None of those two-cell adhesion systems are present in fungi or plants, but the analysis of the first genome sequence of a choanoflagellate, a group of single-celled and colony forming flagellates that are the closest relatives to metazoans, showed that cadherines appeared prior to the metazoan divergence. In fact, up to 23 different cadherins were described in the choanoflagellate Monosiga brevicollis, some of them structurally homologous to some metazoan cadherins (e.g., protocadherin or FAT) and others with completely new domain arrangements.Citation3 However, M. brevicollis does not have an integrin-mediated adhesion system. This leaves the integrin-mediated adhesion and signaling mechanism as one of the key inventions of the metazoan lineage.Citation4,Citation5
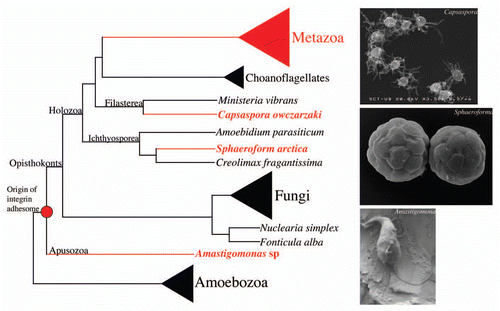
Molecular systematics has shown that choanoflagellates are not the only single-celled metazoan relatives. In fact, the opisthokonts are no longer a game of three (Metazoa, Fungi and choanoflagellates), but rather an eukaryotic clade teaming up with an increasing number of poorly known protist lineages, such as nucleariids, ichthyosporeans, filastereans (namely Capsaspora owczarzaki and Ministeria vibrans), Corallochytrium limacisporum and, very recently, Fonticula alba (see ).Citation6–Citation10 That means those lineages should be taken into account when trying to infer the metazoan and fungi “genetic starter kit”. In this regard, the UNICORN initiative aims to obtain the genome sequence of up to eleven protists lineages closely related to both fungi and metazoans.Citation11 Thanks to this project, the genome sequence of two cythrid fungi (Allomyces macrogynus and Spizellomyces puctatus), one filasterean (C. owczarzaki) and one choanoflagellate (Proterospongia sp.) have been obtained and are publicly available. Moreover, the complete genome of one apusozoan (Amastigomonas sp.), the putative sistergroup of the opisthokontsCitation10,Citation12 has also been sequenced.
This new information allows useful and taxonomically broader comparative genomic analyses, such as the one recently published by Arnau Sebé-Pedrós et al.Citation13 in which the repertoire of the different components of the integrin adhesome in opisthokonts and eukaryotes in general was investigated. The findings of Sebé-Pedrós are indeed unexpected, since a whole integrin-mediated machinery was found in two non-metazoan lineages, the filasterean C. owczarzaki and the apusozoan Amastigomonas sp (see ). That the integrin-mediated complex is present in C. owczarzaki is by itself interesting enough, implying (1) that integrins are not exclusive to Metazoa and (2) that choanoflagellates lost such an important adhesion and signaling mechanisms. However, the most surprising finding is that the genome of the single-celled apusozoan Amastigomonas sp., which is not an opisthokont, also encodes the full repertoire (except for the two associated tyrosine kinases) of the integrin-mediated adhesion complex. These findings have large evolutionary implications. They not only take the integrin exclusivity out of metazoans, but also indicate that fungi (which comprise several species with complex multicellularity) secondarily lost this important cell adhesion and signaling machinery. Moreover, the finding that the origin of integrins is ancient means that the two most important metazoan cell adhesion mechanisms (cadherins and integrins) were already present in pre-metazoan lineages, and they were probably coopted for new functions in metazoans.
Undoubtedly, the presence of integrins in those unicellular organisms opens new and challenging questions. The first one, the actual role that this integrin machinery is playing in those single-celled organisms. Is the integrin machinery involved in sensing the extracellular environment? Such is the case, for example, of cadherins in choanoflagellates, which have been proposed be involved in the response to the extracellular environment, as bacterial prey capture.Citation3 Another question is whether integrins are present in other opisthokont lineages, such as ichthyosporean or nucleariids or Fonticula alba (which are the sister-group to Fungi). In this regard, the genome data of the remaining taxa to be sequenced by the UNICORN initiative (namely F. alba, the ichthyosporeans Sphaeroforma arctica and Amoebidium parasiticum and the free-living filasterean M. vibrans) will surely help unravel the evolutionary history of such important cell adhesion machinery.
Actually, a quick look at the current genome trace data of one ichthyosporean, S. arctica, whose genome is currently being sequenced, shows strong hits to integrins, suggesting that ichthyosporeans most probably also encode the integrin adhesion machinery. That means integrins are found in very different functional contexts, from single-celled amoeba endosymbiont crawlers (such as C. owczarzaki), to marine free-living flagellates (such as Amastigomonas sp.), to colony-forming fish and arthropod parasites (such as the ichthyosporean S. arctica), to fully multicellular eukaryotes (such as Metazoa) (see the ). To us, this capacity to work in such different contexts suggests that integrins most probably had an ancestral role in signaling, for example in sensing the environment to modulate cell physiology and growth. The fact that the IPP signaling module is as ancient as integrins (it is also present in apusozoans, reviewed in ref. Citation13), and that the two tyrosine kinases associated with the integrin machinery (FAK and C-Src) were already present in the common ancestor of C. owczarzaki, choanoflagellates and Metazoa, seem to back up the signaling role of the ancestral integrins. The current cell-extracellular matrix role of the integrin complex in metazoans may have appeared by co-option, although we can not rule out the possibility that integrins in pre-metazoans played a cell adhesion role in, for example, the colony-forming ichthyosporeans. In any case, functional analyses to elucidate the current role of the integrin adhesion system in C. owczarzaki, S. arctica or Amastigomonas sp., will surely be crucial to fully answer these new open questions.
Figures and Tables
Figure 1 Schematic phylogenetic tree of the opisthokonts showing the putative origin of the integrin adhesion system with the current taxon sampling. Relationships are based on both published and unpublished molecular analyses.Citation6–Citation10 Taxa in red are the ones encoding a complete integrin-mediate adhesion machinery. In bold taxa with complete genome sequences. Note that the genome of S. arctica is currently under way, but the trace data shows integrin hits.
Acknowledgements
We thank Lora L. Shadwick and John D.L. Shadwick for editing and critically reading the manuscript. This work was supported by an ICREA contract, an ERC Starting Grant (206883), and a grant (BFU2008-02839/BMC) from MICINN to IR.T. AS.P's salary was supported by a pre-graduate FPU grant from MICINN.
Addendum to:
References
- King N. The unicellular ancestry of animal development. Dev Cell 2004; 7:313 - 325
- Magie CR, Martindale MQ. Cell-cell adhesion in the cnidaria: insights into the evolution of tissue morphogenesis. Biol Bull 2008; 214:218 - 232
- Abedin M, King N. The premetazoan ancestry of cadherins. Science 2008; 319:946 - 948
- King N, Westbrook MJ, Young SL, Kuo A, Abedin M, Chapman, et al. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans. Nature 2008; 451:783 - 788
- Rokas A. The origins of multicellularity and the early history of the genetic toolkit for animal development. Annu Rev Genet 2008; 42:235 - 251
- Ruiz-Trillo I, Inagaki Y, Davis LA, Sperstad S, Landfald B, Roger AJ. Capsaspora owczarzaki is an independent opisthokont lineage. Curr Biol 2004; 14:946 - 947
- Ruiz-Trillo I, Roger AJ, Burger G, Gray MW, Lang BF. A phylogenomic investigation into the origin of metazoa. Mol Biol Evol 2008; 25:664 - 672
- Steenkamp ET, Wright J, Baldauf SL. The protistan origins of animals and fungi. Mol Biol Evol 2006; 23:93 - 106
- Liu Y, Steenkamp ET, Brinkmann H, Forget L, Philippe H, Lang BF. Phylogenomic analyses predict sistergroup relationship of nucleariids and fungi and paraphyly of zygomycetes with significant support. BMC Evol Biol 2009; 9:272
- Brown MW, Spiegel FW, Silberman JD. Phylogeny of the “forgotten” cellular slime mold, Fonticula alba, reveals a key evolutionary branch within Opisthokonta. Mol Biol Evol 2009; 26:2699 - 2709
- Ruiz-Trillo I, Burger G, Holland PW, King N, Lang BF, Roger AJ, et al. The origins of multicellularity: a multi-taxon genome initiative. Trends Genet 2007; 23:113 - 118
- Kim E, Simpson AG, Graham LE. Evolutionary relationships of apusomonads inferred from taxon-rich analyses of 6 nuclear encoded genes. Mol Biol Evol 2006; 23:2455 - 2466
- Sebé-Pedrós A, Roger AJ, Lang FB, King N, Ruiz-Trillo I. Ancient origin of the integrin-mediated adhesion and signaling machinery. Proc Natl Acad Sci USA 2010; 107:10142 - 10147