
Abstract
During cerebral cortical development, post-mitotic neurons exhibit a multi-step migration. The locomotion mode covers most of the neuronal migration path. Although for many decades, locomoting neurons have been known to migrate along radial glial fibers, how the cortical locomoting neurons attach to and migrate along radial glial fibers was largely unknown. We recently reported that N-cadherin is required for cortical neuronal migration in vivo. Knockdown or dominant negative inhibition of N-cadherin results in severe neuronal migration defects. Furthermore, suppression of Rab5-dependent endocytosis increases cell surface levels of N-cadherin and perturbs neuronal migration. We showed here that N-cadherin overexpression, which would mimic Rab5 suppression, weakly suppressed neuronal migration, suggesting that excess N-cadherin also disturbs neuronal migration. Interestingly, however, N-cadherin-knockdown and overexpression in neurons resulted in different morphologies. While N-cadherin-overexpressing cells closely attached to the radial glial fibers similar to control or Rab5-knockdown cells, N-cadherin knockdown weakened the attachment as the average distance between the soma and radial glial fibers was significantly increased. Taken together, these findings suggest that N-cadherin controls the neuronal attachment to radial glial fibers and that N-cadherin-mediated adhesion complexes are reconstituted through Rab GTPases-dependent endocytic pathways to maintain the proper surface N-cadherin level and to promote neuronal migration.
Acknowledgements
We thank Dr. Ozawa for providing us with N-cadherin-expressing vector, and Dr. Ruth T. Yu for helpful comments. We also thank the Core Instrumentation Facility, Keio University School of Medicine for help with Leica SP5 confocal microscopy. This work was supported by Grants-in-Aid from the Ministry of Education, Culture, Sports and Science and Technology, Japan and by grants from the JST PRESTO program and GCOE.
Figures and Tables
Figure 1 Effects on N-cadherin knockdown or overexpression on the locomotion mode of neuronal migration. (A, B, J and K) The morphologies of N-cadherin knockdown or overexpressing locomoting neurons in the intermediate zone of E17 cerebral cortices. The neurons were electroporated with the following plasmids plus pCAG-EGFPCitation19 at E14; Neurons in (A, B, J and K) were electroporated with control scrambled short hairpin RNA (sh-scr4: the negative control vector of pSilencer3.1-H1-neo, Ambion), short hairpin RNA for N-cadherin (NC-sh1023),Citation15 control vector (CAG-MCS2, 10 µg/µl),Citation25 or N-cadherin-expressing vector (CAG-human N-cadherin, 10 µg/µl),Citation23 respectively. Green and red represent immunostaining signals for EGFP or Nestin, a marker for radial glial fibers, respectively. Arrows in (K) indicate branched leading processes. In utero electroporation and immunohistochemistry were performed as described previously in references Citation15 and Citation19. (C and L) The distance between the center of the cell soma of the cells with one thick process in the intermediate zone and the nestin-positive radial glial fiber to which they attach were deduced by Leica SP5 confocal microscopy. Each bar represents the average distance ± SEM. The average distance was significantly increased in N-cadherin-knockdown conditions (sh-scr4: 2.03 ± 0.07 µm, NC-sh1023: 2.92 ± 0.19 µm, p < 0.0001, t-test), whereas it was slightly but significantly decreased in N-cadherin-overexpression (control vector: 1.95 ± 0.04 µm, CAG-N-cadherin: 1.75 ± 0.05 µm, p < 0.01, t-test). (D–I) Cerebral cortices at P0, electroporated with control vector (CAG-MCS2, 10 µg/µl)Citation25 or N-cadherin-expressing vector (CAG-human N-cadherin, 10 µg/µl),Citation23 plus pCAG-EGFP at E14. EGFP fluorescent signals were shown as green in (D and G) or different colors as indicated by the color bar in (E and H). The graphs in (F and I) show the estimation of cell migration, which was carried out by recording fluorescence intensities of EGFP in distinct regions of the cerebral cortices using Leica SP5 software. Each bar represents the mean percentage of relative intensity ± SEM. n = 4 brains. The quantification method was described previously in references Citation14 and Citation19. II–IV, layers II–IV of the cortical plate; V–VI, layers V–VI of the cortical plate; IZ, intermediate zone; WM, white matter; SVZ/VZ, subventricular zone/ventricular zone. Scale bars: (in A) A, B, 10 µm; (in D) D, E, G and H, 200 µm; (in J) J and K, 10 µm.
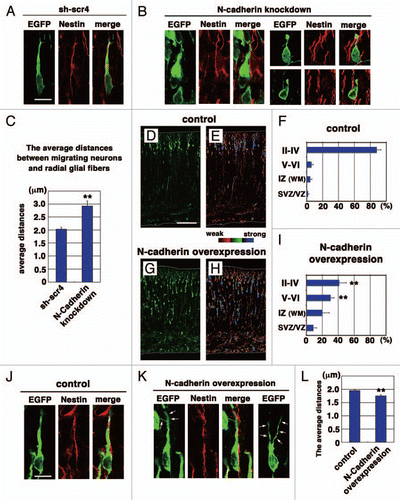
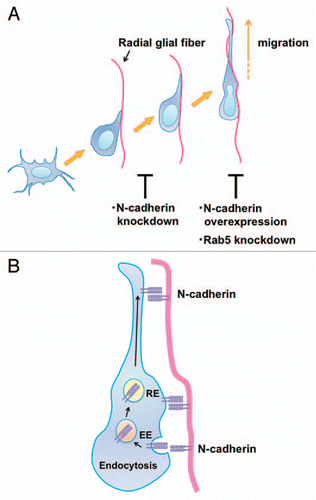
Figure 2 N-cadherin mediates the interaction between migrating neurons and radial glial fibers and radial glial-guided locomotion mode of neuronal migration. (A) Multipolar cells form a thick leading process, retract the other processes and attach to the radial glial fibers in order to change into the locomoting neurons. Knockdown of N-cadherin weakens the interaction between the polarized locomoting neurons and radial glial fibers. Subsequently, the locomoting neurons migrate toward the pial surface along the radial glial fibers. Similar to Rab5 suppression, increased levels of N-cadherin disturb the migration. (B) In the locomoting neurons, a portion of the cell surface N-cadherin is internalized into early endosomes (EE) and recycled back to the plasma membrane via recycling endosomes (RE). The intracellular trafficking of N-cadherin is essential for the piadirected migration of locomoting neurons. N-cadherin endocytosis and recycling are regulated by Rab family small GTPases, Rab5 and Rab11, respectively.
Addendum to: