
Abstract
The Gram-negative opportunistic pathogen Legionella pneumophila replicates in phagocytes within a specific compartment, the Legionella-containing vacuole (LCV). Formation of LCVs is a complex process requiring the bacterial Icm/Dot type IV secretion system and more than 100 translocated effector proteins, which putatively subvert cellular signaling and vesicle trafficking pathways. Phosphoinositide (PI) glycerolipids are pivotal regulators of signal transduction and membrane dynamics in eukaryotes. Recently, a number of Icm/Dot substrates were found to anchor to the LCV membrane by binding to PIs. One of these effectors, SidC, specifically interacts with phosphatidylinositol-4 phosphate [PtdIns(4)P]. Using an antibody against SidC and magnetic beads coupled to a secondary antibody, intact LCVs were purified by immuno-magnetic separation, followed by density centrifugation. This purification strategy is in principle applicable to any pathogen vacuole that carries specific markers. The LCV proteome determined by LC-MS/MS revealed 566 host proteins, including novel components of the endosomal pathway, as well as the early and late secretory trafficking pathways. Thus, LCV formation is a robust process that involves many (functionally redundant) Icm/Dot substrates, as well as the interaction with different host cell vesicle trafficking pathways.
Figures and Tables
Figure 1 Communication of LCVs with endosomal and secretory trafficking pathways. LCVs are decorated with endosomal and secretory markers, which might be acquired by (i) direct fusion with cell organelles, (ii) interaction with transport vesicles shuttling between compartments, or (iii) recruitment from the cytoplasm. L. pneumophila produces two distinct classes of Icm/Dot-translocated effector proteins, which interact with PtdIns(4)P (SidC, SidM)Citation5,Citation18,Citation35 or PtdIns(3)P (LpnE, LidA),Citation15,Citation35 respectively.
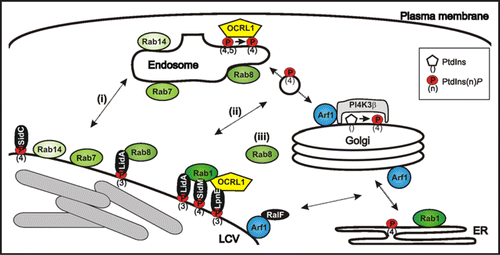
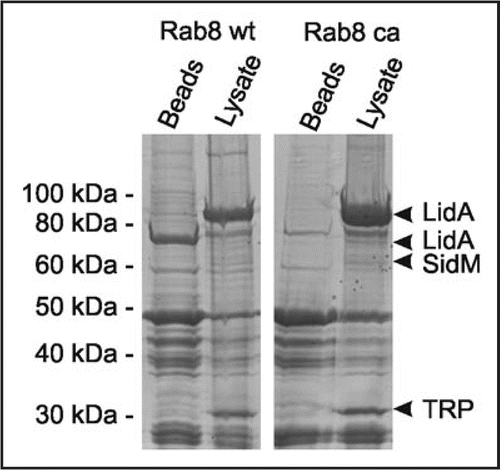
Figure 2 GST-Rab8 interacts with the Icm/Dot substrate LidA in L. pneumophila lysates. GST fusions proteins of Rab8 wild-type (wt) or constitutive active (ca; Rab8Q74LCitation36) were used to precipitate binding partners in L. pneumophila lysates. Protein bands eluting from washed Rab8-coated beads but not from control beads were identified by LC-MS/MS as the Icm/Dot substrate LidA, the Rab1 GEF SidM and a putative tellurite resistance protein (TRP).
Addendum to: