Abstract
The epigenome plays a vital role in helping to maintain and regulate cell functions in all organisms. Alleles with differing epigenetic marks in the same nucleus do not function in isolation but can interact in trans to modify the epigenetic state of one or both alleles. This is particularly evident when two divergent epigenomes come together in a hybrid resulting in thousands of alterations to the methylome. These changes mainly involve the methylation patterns at one allele being changed to resemble the methylation patterns of the other allele, in processes we have termed trans-chromosomal methylation (TCM) and trans-chromosomal demethylation (TCdM). These processes are primarily modulated by siRNAs and the RNA directed DNA methylation pathway. Drawing from other examples of trans-allelic interactions, we describe the process of TCM and TCdM and the effect such changes can have on genome activity. Trans-allelic epigenetic interactions may be a common occurrence in many biological systems.
Bringing together two divergent genomes and epigenomes in the same nucleus has the potential to alter gene expression and the phenotype of an organism. This is evident in plant crop systems where hybrids commonly show increased vigour or heterosis (reviewed in refs Citation1-Citation3). Recent studies have found the epigenome to be a possible contributor to heterosis, especially in intra-species hybrids, where genetic diversity is limited.Citation4-Citation8 In the F1 hybrids, interaction occurs between the epigenomes contributed by the two parental sets of chromosomes leading to altered gene expression.Citation6-Citation8 We have shown that patterns of DNA methylation are altered in intraspecific Arabidopsis hybrids through two processes: trans-chromosomal methylation (TCM) and trans-chromosomal demethylation (TCdM), where the methylation status of one parental allele changes to resemble the levels and patterns of methylation on the other allele ().Citation7 This process is mediated primarily through 24 nt short-interfering RNAs (siRNAs). Our studies have emphasized the extent and complexity of the interactions occurring in the nucleus that govern the activity of the genome and regulation of gene expression. In this review we describe TCM and TCdM and the ways in which they are related to similar trans-allelic epigenetic processes in plants.
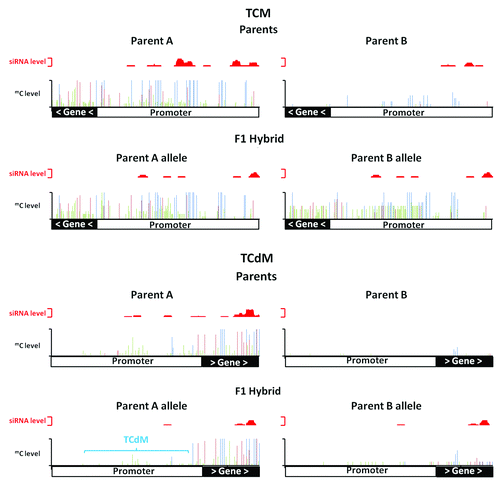
Figure 1. Trans-chromosomal methylation (TCM) and trans-chromosomal demethylation (TCdM). In a TCM event in the hybrid, the unmethylated parental allele (Parent B) increases in methylation to resemble the methylated parental allele (parent A) at locus At3g43340. In a TCdM event in the hybrid, the methylated parental allele (parent A) shows a reduction in methylation (mainly associated with CHH methylation) at a region with reduced levels of siRNAs in the hybrid at locus At2g02660. Methylated CG (blue), CHG (red) and CHH (green) are plotted along the x-axis with levels of methylation and siRNAs plotted on the y-axis. TCM and TCdM correlate with regions producing siRNAs (plotted in red above each methylation graph).
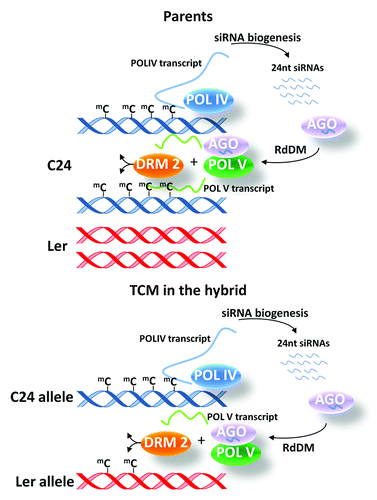
In plants, cytosine methylation occurs in three sequence contexts (CG, CHG and CHH; H = A, C or T), contrasting to animals where methylation is largely restricted to CG dinucleotides (reviewed in ref. Citation9). De novo methylation in plants is generated by the RNA directed DNA methylation (RdDM) pathway where 24 nt siRNAs target regions of the genome for methylation (reviewed in ref. Citation10). The 24 nt siRNAs are produced from a given genomic sequence through the operation of RNA Polymerase IV (POL IV) and then de novo methylation is targeted to homologous genomic sequences by a transcript produced by RNA Polymerase V (POL V), which recruits DNA methyltransferases or, possibly, demethylases (). CG and CHG methylation events are associated with siRNA-independent maintenance pathways, which perpetuate their methylation state through successive cell divisions.Citation11 CHH methylation is maintained almost exclusively by RdDM-dependent de novo methylation after each cell division (reviewed in ref. Citation10). The three contexts differ in abundance, genomic distribution and the extent to which they affect regional activity of the genome.Citation11-Citation13 In the F1 hybrid plants, the majority of sequence segments (loci) associated with siRNA and methylation show levels at the expected mid-parent level (MPV—average of the two parent values; additive inheritance). In fewer than 10% of the loci analyzed, levels of 24 nt siRNA and/or DNA methylation deviated from the MPV (non-additive inheritance). The non-additively expressed siRNAs showed reduced levels at localized regions in the hybrid genome.Citation6 The majority of localized non-additive methylation events have values above the MPV but decreases in methylation are also observed, particularly in the CHH context.Citation7 Throughout the genome non-additive inheritance of siRNAs and methylation in the hybrids occurs most frequently at those loci where parental levels of these epigenetic marks differ.
Figure 2. siRNA-mediated trans chromosomal methylation. The RNA directed DNA methylation pathway plays a key role in TCM and TCdM events. RNA Polymerase IV (POL IV) transcribes a region which following double stranded RNA synthesis and cleavage forms 24 nt siRNAs. Twenty-four nt siRNA are loaded into an ARGONAUTE (AGO) and target regions by an RNA transcript produced by RNA Polymerase V (POL V). This complex that contains additional components can then recruit DOMAIN REARRANGED METHYLTRANSFERASE 2 (DRM2), which methylates the DNA. In hybrids, siRNAs derived from the methylated parental allele target both the methylated parental allele for continued methylation (blue helix) and de novo methylation of the unmethylated parental allele (red helix). After the initial TCM event (illustrated here) the previously unmethylated allele will produce siRNAs and transcripts produced by POL V enabling the newly methylated cytosines to be maintained through subsequent generations.
Non-additive methylation in the hybrids occurs primarily through the TCM or TCdM processes. In TCM, the level of methylation of the low parent allele increases toward that of the stably inherited level of the high parent allele resulting in an overall increase in methylation of all three sequence contexts CG, CHG and CHH (). In TCdM, the methylation level of the high parent allele in the hybrid decreases toward the level of the stably inherited low parent allele, resulting in a loss of methylation mainly associated with the CHH context (). We have estimated that in the Arabidopsis hybrids we examined, TCM and TCdM events occur at several thousand loci. These TCM/TCdM events are not distributed randomly through the genome but occur primarily in those segments of the genome where the 24 nt siRNA levels differ in the parents. This correlation suggests a role for 24 nt siRNAs and the RdDM pathway in directing these trans-allelic events and the alterations to the hybrid methylome. Another mechanism must also exist as TCM and TCdM events can take place independently of siRNAs, but these are primarily associated with CG methylation in gene bodies.Citation7
siRNAs are Effectors in Directing TCM/TCdM Events
The involvement of 24 nt siRNAs in altering genomic methylation patterns through trans-allelic interactions has been described in other plant systems. The Arabidopsis FLOWERING WAGENINGEN A (FWA) gene is involved in regulating flowering and is normally transcriptionally silenced in vegetative tissue by methylation initiated at a set of tandem repeat sequences located in the promoter.Citation14,Citation15 The methylated region includes the repeat elements and the 5′UTR with 24 nt siRNAs being produced from this region. If an unmethylated FWA transgene is introduced into a plant carrying an endogenous methylated FWA allele, the transgene becomes methylated and silenced through a process analogous to TCM in the hybrids.Citation16,Citation17 The transgene remains unmethylated and becomes transcriptionally active if introduced into a plant that lacks the ability to produce siRNAs from the repeat sequences in the FWA promoter. Similarly, if the transgene is introduced into a plant carrying an active unmethylated endogenous fwa gene that still produces siRNAs from the repeat sequence, the transgene is not methylated and becomes transcriptionally active.Citation17,Citation18 Therefore, the presence of both a methylated allele and siRNAs produced from the region are required for trans-allelic methylation in the FWA system. These epigenetic characteristics are seen in many of the loci undergoing TCM in our hybrid system.
The same is true in some examples of paramutation, a system studied in maize for several decades. In paramutation, the transcriptional state of an allele can be altered in a heritable way through the interaction with another allele. The best-studied example is the b1 (Booster 1) locus, which encodes a transcription factor involved in the activation of anthocyanin biosynthesis (reviewed in ref. Citation19). A plant homozygous for the B-I allele at the b1 locus has dark purple pigmentation in its vegetative structures, whereas a plant carrying a B’ allele has only very light purple pigmentation in its vegetative structures. If the two alleles are brought together in the one plant, the B-I allele is epigenetically converted to a B’ state resulting in a light purple phenotype identical to that produced by the B’ parent plant. This again is a trans-interaction that results in TCM between the two alleles mediated through 24 nt siRNAs produced from seven direct repeats located approximately 100 kb upstream of the b1 locus.Citation20-Citation23
In F1 hybrids, TCM events lead to changes in the methylome and potentially gene transcription at thousands of loci across the genome, whereas the FWA gene in Arabidopsis and the b1 locus in maize reflect TCM and gene expression changes at a single locus. Another example of frequent TCM events occurring genome-wide can be seen when wild type plants are crossed to mutant lines that have genome-wide hypomethylation.Citation24-Citation26 The progeny of these crosses following segregation away of the mutant allele, denoted epiRILS (epigenetic Recombinant Inbred Lines), were evaluated over several generations for stability and inheritance of the methylated alleles contributed by the wild type parent and their unmethylated counterparts provided by the hypomethylated mutant. Among the loci, there were some hypomethylated alleles that recovered wild type levels of methylation and others that regained partial levels. In both cases 24 nt siRNAs were associated with these loci. Loci that did not recover methylation at the hypomethylated allele had no associated 24 nt siRNAs. Some loci showed a decrease in methylation of the wild type allele and an associated reduction in siRNA similar to the TCdM events observed in the hybrids.Citation24
siRNAs have been implicated in other trans-allelic events akin to TCM, including the silencing of SP11 alleles involved in the Brassica self-incompatibility system, a system that prevents inbreeding. Among the SP11 alleles, there are some that are dominant in their action relative to other alleles. A dominant allele acts in trans through the production of 24 nt siRNAs from repeats located within the flanking region of the gene to induce TCM in the promoter of a recessive allele preventing its transcription.Citation27 Another example is the methylation of the tryptophan biosynthesis PHOSPHORIBOSYLANTHRANILATE ISOMERASE (PAI) gene family in Arabidopsis, where an inverted repeat between PAI1 and PAI4 produces 24 nt siRNAs that result in TCM and silencing of all four PAI genes in the Arabidopsis Ws ecotype.Citation28,Citation29 When Ws is crossed to the Columbia ecotype, the 24 nt siRNAs produced from the Ws PAI alleles can cause TCM of the Columbia PAI alleles.
In the Arabidopsis hybrids and the examples above, a pool of 24 nt siRNAs contributed by the respective parents interact with both chromosomes. At loci where methylation levels are equivalent between parental alleles, the pool of siRNAs likely associate with both chromosomes but the interaction is not scorable as there is no change in methylation. At loci where methylation levels differ between parental alleles, the association of siRNAs across both chromosomes can result in TCM or TCdM events. Even at loci with differences in methylation between parents, the hybrids often exhibit additive inheritance (i.e., no trans effect) with each allele retaining the methylation level exhibited in the parents. These observations show that siRNAs are necessary but not sufficient to induce TCM and that other factors must be involved.
In the examples of the FWA and b1 genes, the unmethylated actively expressing alleles still produce siRNAs at levels similar to the silenced alleles.Citation17,Citation23 Methylation and silencing of these active alleles can be achieved if siRNA levels are increased through the use of a transgene.Citation15,Citation23 This suggests that the chromatin state of the allele modulates the levels of siRNAs required for methylation and it is likely that this relationship determines the potential for a region to undergo TCM. In the Arabidopsis hybrids, this quantitative relationship between siRNAs and the chromatin state of the recipient allele may be the reason why in situations where parental alleles differ in siRNA and methylation levels we can observe either TCM, TCdM or additive inheritance (reviewed in ref. Citation30).
siRNA-Independent TCM
Although the 24 nt siRNAs are involved in altering methylation patterns in hybrids, we identified some TCM events that were independent of siRNAs, mainly consisting of CG methylation within gene bodies. Unlike those TCM events where siRNAs are mediators for de novo methylation in trans, a mechanism for siRNA-independent TCM is not obvious. De novo DNA methylation can occur without involving siRNAs, through interactions between chromatin marks such as H3K9 dimethylation and cytosine methyltransferases.Citation31 However, this process occurs in cis and would have to be transferred to the other parental allele in the hybrid for TCM to occur. An alternative possibility is that, since CG gene body methylation is associated with gene expression,Citation32,Citation33 the siRNA independent TCM events could be generated through changes in allelic activity driven by differences between the parental regulatory components now combined in the hybrid.
Inheritance of TCM and TCdM Generated Methylation Patterns
The epigenetic change of state between the B' and B-I alleles at the b1 locus shows full penetrance in the F1 offspring and is stably inherited through successive cell and sexual generations (reviewed in ref. Citation19). There may be a different situation in the Arabidopsis hybrids since the heterotic phenotype of larger biomass decreases in subsequent generations. This could imply that the epigenetic changes that differentiate the F1 hybrid from the parental chromosomal states may be altered in the F2 and later generations. The F1 Arabidopsis hybrids show disparity in the penetrance of TCM and TCdM at different loci, with some recipient alleles completely matching the methylation profile of the donor allele, while at other loci the recipient allele shows only modest changes in methylation.Citation7 Low levels of TCM-derived methylation in the F1 hybrid may increase in subsequent generations as occurs at the PAI loci and many of the alleles found in epiRILs.Citation26,Citation28 Conversely, alleles that gain methylation through TCM could revert back to an unmethylated state in subsequent generation as occurs in some loci undergoing paramutation.Citation34 At this stage, we have no data on the inheritance of TCM or TCdM events in the F2 and later generations of the hybrid.
TCM in Mammals
Trans-allelic epigenetic interactions also occur in mammals (reviewed in refs. Citation34 and Citation35). The most similar example to siRNA-dependent TCM in plants occurs at the RAS-protein-specific guanine nucleotide-releasing factor (Rasgrf1) locus found on chromosome 9 in mice (reviewed in refs. Citation34 and Citation35). Rasgrf1 is an imprinted gene with monoallelic expression of the paternal allele modulated through methylation of a regulatory element in the parental allele, which blocks binding of the CTCF transcriptional repressor (reviewed in ref. Citation36). TCM of this CTCF binding element occurs through a non-coding RNA overlapping this region being targeted by piwi-interacting RNAs (piRNAs) originating from chromosome 7. This process recruits the DNMT3A methyltransferase leading to methylation of the paternal Rasgrf1 allele. Mutants that are unable to produce piRNAs from the chromosome 7 locus fail to undergo TCM at the paternal Rasgrf1 allele.Citation37
Final Thoughts
It is clear that the epigenetic states of two different genomes in the same nucleus do not function in isolation but interact to produce altered epigenomes. The hybrid system presents a powerful opportunity to understand the generation of new epigenetic states that may produce significant biological outcomes such as heterosis. Our findings that TCM and TCdM events occur at several thousand loci in a system as common as hybrids indicates that cross-talk between epialleles is a regular biological occurrence that can affect gene activity and ultimately development. The relationship between 24 nt siRNAs and DNA methylation in influencing the transcriptional activity of a gene is not well understood, but TCM and TCdM events frequently occur in control regions of genes in the hybrid. In the hybrids, we have only detailed a few examples of the relationship between methylation status of a gene and its flanking regions with its transcriptional activity. More needs to be done at a single gene level to clarify the relationship between these epigenetic systems and gene activity. Other components of the epigenome, such as chemical modifications of histones, need to be investigated in hybrids as these epigenetic marks play a key role in regulating and silencing of genes and may influence the ability of siRNA-mediated TCM or TCdM events to occur at individual loci. With a fuller understanding of these relationships, it may be ultimately possible to make epigenome constructs to maximize and/or optimize particular gene activities or gene pathway activities. Additionally, it may be possible to gain particular phenotypes through manipulation of either the epigenome or the genome and perhaps, most significantly, of the way in which they interact.
Related Research Data
References
- Lippman ZB, Zamir D. Heterosis: revisiting the magic. Trends Genet 2007; 23:60 - 6; http://dx.doi.org/10.1016/j.tig.2006.12.006; PMID: 17188398
- Hochholdinger F, Hoecker N. Towards the molecular basis of heterosis. Trends Plant Sci 2007; 12:427 - 32; http://dx.doi.org/10.1016/j.tplants.2007.08.005; PMID: 17720610
- Birchler JA, Yao H, Chudalayandi S, Vaiman D, Veitia RA. Heterosis. Plant Cell 2010; 22:2105 - 12; http://dx.doi.org/10.1105/tpc.110.076133; PMID: 20622146
- He GM, Zhu XP, Elling AA, Chen LB, Wang XF, Guo L, et al. Global epigenetic and transcriptional trends among two rice subspecies and their reciprocal hybrids. Plant Cell 2010; 22:17 - 33; http://dx.doi.org/10.1105/tpc.109.072041; PMID: 20086188
- Ha M, Lu J, Tian L, Ramachandran V, Kasschau KD, Chapman EJ, et al. Small RNAs serve as a genetic buffer against genomic shock in Arabidopsis interspecific hybrids and allopolyploids. Proc Natl Acad Sci U S A 2009; 106:17835 - 40; http://dx.doi.org/10.1073/pnas.0907003106; PMID: 19805056
- Groszmann M, Greaves IK, Albertyn ZI, Scofield GN, Peacock WJ, Dennis ES. Changes in 24-nt siRNA levels in Arabidopsis hybrids suggest an epigenetic contribution to hybrid vigor. Proc Natl Acad Sci U S A 2011; 108:2617 - 22; http://dx.doi.org/10.1073/pnas.1019217108; PMID: 21266545
- Greaves IK, Groszmann M, Ying H, Taylor JM, Peacock WJ, Dennis ES. Trans chromosomal methylation in Arabidopsis hybrids. Proc Natl Acad Sci U S A 2012; 109:3570 - 5; http://dx.doi.org/10.1073/pnas.1201043109; PMID: 22331882
- Shen H, He H, Li J, Chen W, Wang X, Guo L, et al. Genome-wide analysis of DNA methylation and gene expression changes in two Arabidopsis ecotypes and their reciprocal hybrids. Plant Cell 2012; 24:875 - 92; http://dx.doi.org/10.1105/tpc.111.094870; PMID: 22438023
- He X-J, Chen T, Zhu J-K. Regulation and function of DNA methylation in plants and animals. Cell Res 2011; 21:442 - 65; http://dx.doi.org/10.1038/cr.2011.23; PMID: 21321601
- Matzke M, Kanno T, Huettel B, Daxinger L, Matzke AJM. Targets of RNA-directed DNA methylation. Curr Opin Plant Biol 2007; 10:512 - 9; http://dx.doi.org/10.1016/j.pbi.2007.06.007; PMID: 17702644
- Law JA, Jacobsen SE. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat Rev Genet 2010; 11:204 - 20; http://dx.doi.org/10.1038/nrg2719; PMID: 20142834
- Lister R, O’Malley RC, Tonti-Filippini J, Gregory BD, Berry CC, Millar AH, et al. Highly integrated single-base resolution maps of the epigenome in Arabidopsis. Cell 2008; 133:523 - 36; http://dx.doi.org/10.1016/j.cell.2008.03.029; PMID: 18423832
- Cokus SJ, Feng SH, Zhang XY, Chen ZG, Merriman B, Haudenschild CD, et al. Shotgun bisulphite sequencing of the Arabidopsis genome reveals DNA methylation patterning. Nature 2008; 452:215 - 9; http://dx.doi.org/10.1038/nature06745; PMID: 18278030
- Soppe WJJ, Jacobsen SE, Alonso-Blanco C, Jackson JP, Kakutani T, Koornneef M, et al. The late flowering phenotype of fwa mutants is caused by gain-of-function epigenetic alleles of a homeodomain gene. Mol Cell 2000; 6:791 - 802; http://dx.doi.org/10.1016/S1097-2765(05)00090-0; PMID: 11090618
- Kinoshita Y, Saze H, Kinoshita T, Miura A, Soppe WJJ, Koornneef M, et al. Control of FWA gene silencing in Arabidopsis thaliana by SINE-related direct repeats. Plant J 2007; 49:38 - 45; http://dx.doi.org/10.1111/j.1365-313X.2006.02936.x; PMID: 17144899
- Cao XF, Jacobsen SE. Role of the arabidopsis DRM methyltransferases in de novo DNA methylation and gene silencing. Curr Biol 2002; 12:1138 - 44; http://dx.doi.org/10.1016/S0960-9822(02)00925-9; PMID: 12121623
- Chan SWL, Zhang X, Bernatavichute YV, Jacobsen SE. Two-step recruitment of RNA-directed DNA methylation to tandem repeats. PLoS Biol 2006; 4:e363; http://dx.doi.org/10.1371/journal.pbio.0040363; PMID: 17105345
- Chan SWL, Zilberman D, Xie ZX, Johansen LK, Carrington JC, Jacobsen SE. RNA silencing genes control de novo DNA methylation. Science 2004; 303:1336; http://dx.doi.org/10.1126/science.1095989; PMID: 14988555
- Chandler VL. Paramutation’s properties and puzzles. Science 2010; 330:628 - 9; http://dx.doi.org/10.1126/science.1191044; PMID: 21030647
- Stam M, Belele C, Dorweiler JE, Chandler VL. Differential chromatin structure within a tandem array 100 kb upstream of the maize b1 locus is associated with paramutation. Genes Dev 2002; 16:1906 - 18; http://dx.doi.org/10.1101/gad.1006702; PMID: 12154122
- Haring M, Bader R, Louwers M, Schwabe A, van Driel R, Stam M. The role of DNA methylation, nucleosome occupancy and histone modifications in paramutation. Plant J 2010; 63:366 - 78; http://dx.doi.org/10.1111/j.1365-313X.2010.04245.x; PMID: 20444233
- Alleman M, Sidorenko L, McGinnis K, Seshadri V, Dorweiler JE, White J, et al. An RNA-dependent RNA polymerase is required for paramutation in maize. Nature 2006; 442:295 - 8; http://dx.doi.org/10.1038/nature04884; PMID: 16855589
- Arteaga-Vazquez M, Sidorenko L, Rabanal FA, Shrivistava R, Nobuta K, Green PJ, et al. RNA-mediated trans-communication can establish paramutation at the b1 locus in maize. Proc Natl Acad Sci U S A 2010; 107:12986 - 91; http://dx.doi.org/10.1073/pnas.1007972107; PMID: 20616013
- Reinders J, Wulff BBH, Mirouze M, Marí-Ordóñez A, Dapp M, Rozhon W, et al. Compromised stability of DNA methylation and transposon immobilization in mosaic Arabidopsis epigenomes. Genes Dev 2009; 23:939 - 50; http://dx.doi.org/10.1101/gad.524609; PMID: 19390088
- Johannes F, Porcher E, Teixeira FK, Saliba-Colombani V, Simon M, Agier N, et al. Assessing the impact of transgenerational epigenetic variation on complex traits. PLoS Genet 2009; 5:e1000530; http://dx.doi.org/10.1371/journal.pgen.1000530; PMID: 19557164
- Teixeira FK, Heredia F, Sarazin A, Roudier F, Boccara M, Ciaudo C, et al. A role for RNAi in the selective correction of DNA methylation defects. Science 2009; 323:1600 - 4; http://dx.doi.org/10.1126/science.1165313; PMID: 19179494
- Tarutani Y, Shiba H, Iwano M, Kakizaki T, Suzuki G, Watanabe M, et al. Trans-acting small RNA determines dominance relationships in Brassica self-incompatibility. Nature 2010; 466:983 - 6; http://dx.doi.org/10.1038/nature09308; PMID: 20725042
- Luff B, Pawlowski L, Bender J. An inverted repeat triggers cytosine methylation of identical sequences in Arabidopsis. Mol Cell 1999; 3:505 - 11; http://dx.doi.org/10.1016/S1097-2765(00)80478-5; PMID: 10230403
- Enke RA, Dong Z, Bender J. Small RNAs prevent transcription-coupled loss of histone H3 lysine 9 methylation in Arabidopsis thaliana. PLoS Genet 2011; 7:e1002350; http://dx.doi.org/10.1371/journal.pgen.1002350; PMID: 22046144
- Groszmann M, Greaves IK, Albert N, Fujimoto R, Helliwell CA, Dennis ES, et al. Epigenetics in plants-vernalisation and hybrid vigour. Biochim Biophys Acta 2011; 1809:427 - 37; PMID: 21459171
- Sasaki TKA, Kobayashi A, Saze H, Kakutani T. RNAi-independent de novo DNA methylation revealed in Arabidopsis mutants of chromatin remodeling gene DDM1. Plant J 2012; 70:750 - 8; http://dx.doi.org/10.1111/j.1365-313X.2012.04911.x; PMID: 22269081
- Zhang XY, Yazaki J, Sundaresan A, Cokus S, Chan SWL, Chen HM, et al. Genome-wide high-resolution mapping and functional analysis of DNA methylation in arabidopsis. Cell 2006; 126:1189 - 201; http://dx.doi.org/10.1016/j.cell.2006.08.003; PMID: 16949657
- Zilberman D, Gehring M, Tran RK, Ballinger T, Henikoff S. Genome-wide analysis of Arabidopsis thaliana DNA methylation uncovers an interdependence between methylation and transcription. Nat Genet 2007; 39:61 - 9; http://dx.doi.org/10.1038/ng1929; PMID: 17128275
- Pilu R. Paramutation: just a curiosity or fine tuning of gene expression in the next generation?. Curr Genomics 2011; 12:298 - 306; http://dx.doi.org/10.2174/138920211795860099; PMID: 22131875
- Daxinger L, Whitelaw E. Transgenerational epigenetic inheritance: more questions than answers. Genome Res 2010; 20:1623 - 8; http://dx.doi.org/10.1101/gr.106138.110; PMID: 21041414
- Ferguson-Smith AC. Genomic imprinting: the emergence of an epigenetic paradigm. Nat Rev Genet 2011; 12:565 - 75; http://dx.doi.org/10.1038/nrg3032; PMID: 21765458
- Watanabe T, Tomizawa S-i, Mitsuya K, Totoki Y, Yamamoto Y, Kuramochi-Miyagawa S, et al. Role for piRNAs and noncoding RNA in de novo DNA methylation of the imprinted mouse Rasgrf1 locus. Science 2011; 332:848 - 52; http://dx.doi.org/10.1126/science.1203919; PMID: 21566194