Abstract
Diagnosis of bacterial sepsis in preterm neonates can be difficult when using serum markers that rely on physiological changes because these changes may not necessarily be the result of bacterial infections alone. This retrospective investigation explores the potential use of the DNA methylation pattern of CpG sites in the promoter region of the calcitonin-related polypeptide α (CALCA) gene as an epigenetic biomarker for bacterial sepsis in preterm newborns. Four novel changes in the DNA methylation of eight CpG sites were detected in this gene and are present only in neonates with bacterial sepsis: (1) partial methylation at -769 CpG in gram-negative or gram-positive early onset sepsis (EOS) and late onset sepsis (LOS) episodes; (2) demethylation of 8 CpGs in gram-negative EOS followed by LOS (ELS) and in gram-negative EOS; (3) demethylation of 7 CpGs in gram-positive ELS and gram-positive EOS; (4) -771 C:G > T:A; 5′ de novo -778 CpG mutation on both alleles in EOS. These changes were not detected in birth weight and gestational age matched controls or in newborns with isolated infections. Our results indicate that the DNA methylation pattern of the promoter region of the CALCA gene varies in different types of bacterial preterm sepsis, thus suggesting a potential use as an epigenetic biomarker. A prospective confirmation of these results is essential.
Keywords: :
Introduction
Sepsis is a generalized inflammatory response induced by bloodstream pathogens.Citation1 World Health Organization data show that sepsis accounts for 1.1 million neonatal deaths per year worldwide.Citation2-Citation4 Newborn sepsis is defined by the observation of clinical signs and by blood cultures and can be diagnosed within the first 72 h of life (early onset sepsis; EOS) or after 72 h of life (late onset sepsis; LOS).Citation5 Preterm children with low birth weight (BW < 2500 g) and very low birth weight (BW < 1500 g) are at high risk of acquiring sepsis.Citation2,Citation3,Citation6,Citation7 In term and preterm infants, the most common pathogens associated with EOS are Group B Streptococci (GBS) and Escherichia coli (E. coli). E. coli is frequently seen in preterm infants.Citation8,Citation9 In LOS, the most frequent pathogen is Coagulase-negative Staphylococci (CoNS).Citation5,Citation6,Citation10
The “gold standard” for the diagnosis of bacterial sepsis is the growth of an organism in a blood culture, though only approximately 20% of the cases reveal positive cultures (“culture-positive”). Serum markers for the diagnosis of sepsis are often insufficient, since they rely on physiological changes that may not be the result of the bacterial infection alone and may overlap with those of a sepsis.Citation5,Citation11,Citation12 DNA methylation-based biomarkers, on the other hand, have an advantage over protein expression-based laboratory sepsis markers because they are, in most cases, chemically stable and cannot be modified by physiological conditions that are not related to sepsis.Citation13 DNA methylation changes are associated with the CpG dinucleotides of the gene body and promoter region.Citation14 The 5′ flanking promoter region of the CALCA gene has a large CpG island and has been postulated to contain microbial infection-specific response elements that regulate the transcription of procalcitonin (PCT) during a bacterial infection.Citation15
This makes the promoter region of the CALCA gene a suitable candidate for DNA methylation changes with functional consequences. CALCA is a gene that codes for two alternatively spliced products, calcitonin and calcitonin gene related peptide (α-CGRP), both of which are cleaved from larger pro-peptides PCT and proCGRP, respectively. The CALCA gene is located on chromosome 11 and contains six exons. Exon 1 is non-coding and exon 2 codes for the signal sequences.Citation15 PCT is an acute phase protein whose serum concentration increases in response to a bacterial infection. Bacterial toxins or cytokines released in response to bacterial infections activate the production of PCT from leukocytes and peripheral blood mononuclear cells.Citation16-Citation18
The purpose of this investigation was to retrospectively evaluate the methylation status of CpG sites in the promoter region of the CALCA gene in bacterial sepsis in preterm neonates, using a DNA methylation screening method that utilizes only very small amounts of blood (10 uL).
Results
During the study period, clinical data and blood samples from 49 preterm babies were collected. Babies were distributed into five groups according to the incidence of sepsis during hospitalization. Sixteen of these babies were never suspected of having sepsis until dismissal and served as controls, 10 showed EOS (EOS group), 8 showed EOS with a subsequent episode of LOS (ELS group), 10 showed LOS (LOS group), and 5 babies showed isolated infections (infection group). The clinical characteristics of each group are described in Table S1). A Kruskal Wallis analysis of the study groups resulted in comparable distributions of birth weight and gestational age (GA) across the 5 groups (birth weight, P = 0.359; GA, P = 0.087).
Analysis of DNA methylation status of the promoter regions of 8 genes encoding proteins activated during sepsis, Toll-like receptor 2 (TLR2), Toll-like receptor 4 (TLR4), Myeloid differentiation primary response (MyD88), Interleukin-6 (IL6), Interleukin-6 receptor (IL6R), interleukin-6 signal transducer (gp130), Interleukin-8 (IL8), and C-reactive protein (CRP),Citation19 did not show any differences when comparing sepsis babies to birth weight and gestational age matched controls (results not shown). Altered DNA methylation status was found only in the CALCA gene when comparing sepsis babies to birth weight and gestational age matched controls (–, and ; Figs. S1–S3; Tables S1 and S2).
Figure 1.CALCA gene: schematic representation of the 5′ untranslated promoter region indicating the CpG dinucleotides at the positions -796, -771, -769, -739, -733, -728, -719, -716, and the structural region indicating the exons 2–6.
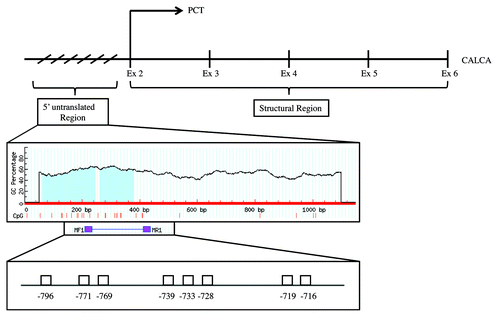
Figure 3. Representative results of the sequencing of CpG dinucleotides at the positions -796, -771, -769, -739, -733, -728, -719, -716 in the 5' untranslated promoter region of the CALCA gene in preterm neonates with bacterial sepsis and birth weight and age matched controls respectively.(A) A -771 C:G > T:A; 5′de novo -778 CpG mutation on both alleles in EOS (day 3). (B) A total methylation of CpG at position -771 and a demethylation at position -778 in birth weight and age matched control
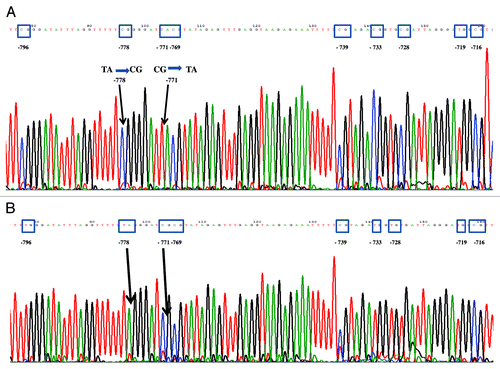
Table 1. Clinical features and DNA methylation changes in preterm newborns affected by EOS
Table 2. Clinical features and DNA methylation changes in preterm newborns affected by ELS (A) and isolated infections (B) respectively
A methylation screening method was established for the screening of the 228-bp promoter region of the CALCA gene (). A representative result (Fig. S3) showed a temperature shift of the EOS (day 3) methylation peak to the right and a temperature shift of the ELS (day 3) methylation peak to the left. A sample collected prior to a LOS episode (day 3) and a control sample (day 3) showed overlapping methylation peaks.
We studied the methylation status of 8 CpG sites (positions -796, -771, -769, -739, -733, -728, -719, and -716) in the promoter region of the CALCA gene () by sequencing 10 EOS, 8 ELS, 10 LOS, 16 controls and 5 isolated infections. A representative result of the sequencing is shown in and , and in Figures S1 and S2.
Figure 2. Representative results of the sequencing of CpG dinucleotides at the positions -796, -771, -769, -739, -733, -728, -719, -716 in the 5' untranslated promoter region of the CALCA gene in preterm neonates with bacterial sepsis and birth weight and age matched controls respectively. (A) Partial methylation of CpG at position -769 in gram-positive EOS (day 1). (B) Total methylation of CpG at position -769 in birth weight and age matched control.
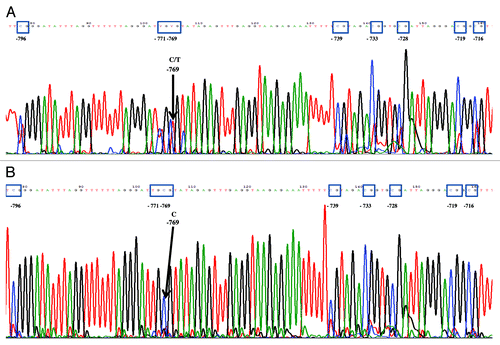
Four novel changes in the DNA methylation pattern of the CALCA promoter were identified: (1) partial methylation at -769 CpG (); (2) Demethylation of 8 CpGs (Fig. S1A); (3) Demethylation of 7 CpGs (Fig. S2A) and; (4) -771 C:G > T:A; 5′de novo -778 CpG mutation on both alleles (). The clinical and epigenetic characteristics of patients and controls were summarized in and , and in Tables S1 and S2.
shows the methylation status of 10 EOS samples (EOS 1-EOS 10) and 16 controls. Five samples (EOS 1, 2, 3, 5, 8) showed partial methylation at -769 CpG, one sample (EOS 7) showed demethylation of 7 CpGs, two samples (EOS 6, 9) showed demethylation of 8 CpGs, and one sample (EOS 10) showed -771 C:G > T:A; 5′de novo -778 CpG mutation on both alleles. None of the 16 controls showed any of the four described methylation changes.
shows the methylation status of 8 ELS samples (ELS 1-ELS 8) and 5 samples with an isolated infection (I1-I5). One sample (ELS 1.2) presented partial methylation at -769 CpG; five samples (ELS 2, 3, 5, 7, 8) showed demethylation of 7 CpGs, and one sample (ELS 1.1) showed demethylation of 8 CpGs. Five samples with an isolated infection (I1-I5) did not show any of the four DNA methylation changes.
Table S2 shows the methylation status of 10 LOS samples (LOS 1-LOS 10). Three samples (LOS 1, 2, 10) showed partial methylation at -769 CpG; seven LOS samples (3, 4, 5, 6, 7, 8, 9) did not show any of the four DNA methylation changes.
Discussion
Changes in the DNA methylation (partial methylation at -769 CpG, demethylation of 8 CpGs, demethylation of 7 CpGs, and -771 C:G > T:A; 5′de novo -778 CpG mutation on both alleles) were detected in the promoter region of the CALCA gene in preterm neonates with bacterial sepsis. These changes seem to be a specific feature of the CALCA gene, as analysis of other genes (TLR2, TLR4, MyD88, IL6, IL6R, gp130, IL8, and CRP) that are also activated during sepsis did not show these DNA methylation changes in their promoter region.
In EOS, partial methylation at -769 CpG did not seem to distinguish gram-positive from gram-negative bacterial infection and was detected in culture-positive and culture-negative samples. Samples that were suspicious for sepsis but were culture-negative presented either amniotic fluid infection or preterm premature rupture of membranes during pregnancy. We speculate that partial methylation at -769 CpG occurs as a result of a bacterial infection in the uterus of the mother. This interpretation is consistent with the literature that shows that a maternal infection by Campylobacter rectus causes changes in the methylation status of the Insulin-like growth factors 2 (IGF2) gene promoter in murine placental tissue, suggesting an epigenetic effect.Citation20
Results using serum markers for diagnosing sepsis are often similar for gram-negative and gram-positive infections.Citation21 We detected specific demethylation of 8 CpGs in gram-negative EOS. In contrast, specific demethylation of 7 CpGs in gram-positive EOS seems to be indicative of an isolated gram-positive infection. Demethylation of 8 CpGs or 7 CpGs was detected in culture-positive samples only. Polymicrobial cultures of gram-negative/gram-positive or fungal/gram-positive did not show demethylation of those 7 CpGs or 8 CpGs, which suggests reduced sensitivity of CALCA gene methylation status for either gram-negative or gram-positive bacteria in a mixed culture.
The relationship of partial methylation at -769 CpG, demethylation of 8 CpGs, demethylation of 7 CpGs, and -771 C:G > T:A; 5′de novo -778 CpG mutation on both alleles to a bacterial infection is underlined by the fact that none of the controls showed these methylation changes, thus implying that they could serve as classical epigenetic infection markers. Moreover, samples collected prior to a LOS episode showed none of these methylation changes, while blood samples collected during LOS episodes showed partial methylation at -769 CpG.
Nevertheless, one LOS sample that was collected during a LOS episode showed no partial methylation at -769 CpG. This particular patient suffered from a mixed bacteria/candida infection under immunosuppressing therapy. We speculate that immunosuppressing therapy could have influenced the partial methylation at -769 CpG. This interpretation is consistent with the literature that shows that induced DNA methylation in gastric mucosa is reduced upon treatment with the immunosuppressor cyclosporin.Citation22 Additionally, Montagna et al. reported that PCT is a weak marker for fungal blood stream infections.Citation23
The possibility of therapy monitoring was illustrated by blood samples collected on two occasions from a patient during a LOS episode. These samples showed partial methylation at -769 CpG. A blood sample collected on a third occasion, with the patient in remission, did not show partial methylation at -769 CpG.
One additional finding worth mentioning was that patients with isolated bacterial infections (pneumonia, urinary tract infections) and fungal infections did not show methylation changes in the promoter region of the CALCA gene. All these observations are indicative that the epigenetic changes in the CALCA gene are directly or indirectly related to bacterial sepsis. Moreover, it seems that different pathogen constellations lead to different methylation patterns.
We speculate that the -771 C:G > T:A; 5′de novo -778 CpG mutation on both alleles in an EOS culture-negative sample from an extremely premature infant (28 weeks GA, 560 g) who died 6 d after birth due to septic shock related multi organ failure, may have increased the susceptibility for bacterial sepsis. Positions -769 CpG and -771 CpG are within a consensus binding site (ACCGCGCA) for specificity protein 1 (Sp1), which suggests putative functional consequence of partial methylation at -769 CpG and mutation at -771 CpG. Sp1 has been described to regulate gene expression.Citation24,Citation25 CpG sites -771 and -778 could be mutation hotspots in the promoter region of the CALCA gene.Citation26
The mechanisms responsible for the change in DNA methylation pattern at the promoter region of the CALCA gene in bacterial sepsis of preterm neonates are unknown. However, evidence in the literature suggests that DNA methylation status could be altered by bacterial infection or toxins.Citation27,Citation28 We have established a screening method that distinguishes different sepsis episodes and utilizes small sample volumes (10 μl). This is an advantage over existing methods because the biggest problem for the confirmation of sepsis in newborns, especially premature newborns, is that only very small amounts of blood can be obtained for analysis.Citation29
Conclusions
We could demonstrate that epigenetic analysis of a gene encoding an acute phase protein involved in infection response may have the potential to serve as a useful biomarker for sepsis in the future. In summary, the DNA methylation status of the promoter region of the CALCA gene in preterm infants with bacterial sepsis is different from the methylation status in infants that show no clinical features of systemic infection. Additionally, the changes seem to be specific to the biology of the affecting pathogens or their specific host response. A prospective study using a larger sample size is needed in order to confirm these initial results.
Materials and Methods
Study design and inclusion criteria
Retrospective analysis of preterm neonates (n = 49; 380–3600 g, GA = 23.00–35.86 weeks) in the Austrian newborn screening register of the pediatric clinic of the University of Vienna medical school was performed with blood spots. Medical records of preterm neonates born prior to 36 weeks of GA hospitalized at two neonatal intensive care unit (NICU) and one intermediate care unit (IMC) located at the department of Pediatrics and Adolescence Medicine of the Medical University of Vienna were monitored for the onset of sepsis or non-systemic infections from 12/2010 to 05/2013. The inclusion criteria for sepsis were defined as clinical suspected sepsis and/or positive blood culture result. Clinical suspicion of sepsis was defined as: IL 8 > 200 mg/dL and/or CRP > 1.5 mg/dL, immature to total neutrophil (I:T) ratio ≥ 0.2, clinical signs at onset of sepsis, and antibiotic treatment ≥5 d. Sepsis was subcategorized into 3 groups: Group 1; EOS, Group 2; ELS and Group 3; LOS. EOS was defined as the onset of clinical symptoms and/or a proven blood culture within the first 72 h of life, ELS indicates the occurrence of a first septical episode within the first 72h of life (culture proven EOS or clinically suspected EOS) followed by one or more LOS episodes until discharge. The time between the septical episodes of EOS and LOS varies from patient to patient. LOS was defined as the onset of symptoms consistent with sepsis later than 72 h of life.Citation5 Additional patients with no evidence for systemic or isolated infection until dismissal from NICU/IMC were selected as gestational age (GA) and birth weight matched controls. Of these patients who were selected according to the inclusion criteria, three 3.1 mm disks of the corresponding newborn screening dried bloodspot filter card (DBS) have been archived for the DNA methylation analysis. DBS cards were stored at room temperature until analysis. For some patients more than 1 DBS card was available for analysis, due to repeat measurements in the routine newborn-screening program. The ethics committee of the Medical University of Vienna approved this investigation (EK 789/2010).
DNA extraction
DNA was extracted from 3 dried blood spot filter paper samples (d = 3.1 mm, 10 µl whole blood) using a QIAmp DNA Blood Mini Kit (Qiagen). DNA was eluted in 45 ul buffer.
Bisulphite modification
Epitect Bisulphite Kit (Qiagen) was used for the bisulphite modification of DNA. A protocol that enables a bisulphite conversion of unmethylated cytosines in DNA from low-concentrated solutions was used and 1–500 ng DNA in a maximum volume of 40 μl was processed. Bisulphite modified DNA was eluted in 20 μl buffer. A positive methylated and bisulfite converted human DNA (Qiagen) was used as a control.
Methylation specific PCR- High resolution melting
Methylation specific PCR (MSP) and high resolution melting (HRM) were performed sequentially on a Rotor Gene 6000 (Corbett Research) using EpiTect HRM PCR Kit (Qiagen). MSP was performed in a mixture containing 5 μl EpiTect HRM Master Mix (EpiTect PCR buffer with EvaGreen dye, HotstarTaq Plus DNA Polymerase, dNTP mix), 1 μl (10 pmol each) of the primer pair covering 252 bp of the 5′ untranslated region of the promoter region, 1ul Mg2+ (24 mM), and 3ul bisulphite modified DNA (5–10ng) or 1 ul positive methylated and bisulfite converted human DNA control in a reaction volume of 10 ul. Amplification was as follows: denaturation at 95 °C, 6 min; 10 cycles of touchdown cycling at 95 °C, 10 sec; 65 °C -0.5 °C each cycle, 30 sec ; 72 °C, 10 sec; and 40 cycles at 95 °C, 10 sec; 60.5 °C, 30 sec; 72 °C, 10 sec; After cycling, heteroduplexes were built at 95 °C, 30 sec; 50 °C, 30 sec and HRM analysis was subsequently performed at a temperature ramping from 60–90 °C, rising 0.1 °C/2 sec as recommended by the manufacturer.
Primers
Primers specific for methylated DNA only were designed using the Methprimer softwareCitation30 which integrates the principles outlined by Wojdacz et al.Citation31 The primers used to amplify bisulphite treated DNA were:
CALCA intr1MgcFw 5′CGCCCGCCGC GCCCCGCGCC CGTCTTGCGG ATTTTAGTTT TATTTTTTC 3′ and CALCA intr1MRev 5′AACTACTTAT CCCTCAATCG TCGTA 3′
GC-clamp: CGCCCGCCGC GCCCCGCGCC CGTC (24bp)
The amplified region is the 5′ untranslated region of CALCA (OMIM 114130) and corresponds to the nucleotides -796 to -716 (ATG is +1) that encompasses eight CpG dinucleotides.
Sequencing
After MSP-HRM, DNA fragments (252bp) were eluted from 0.2% agarose gel and purified with a PCR cleanup kit (Qiagen). Sequencing was performed by an ABI 370 sequencer (VBC-Biotech) in a non-quantitative manner to confirm the presence or absence of DNA methylation changes in samples analyzed.
| Abbreviations: | ||
| CALCA | = | calcitonin-related polypeptide alpha |
| CpG | = | cytosine phosphodiester bond guanine |
| MSP | = | methylation specific PCR |
| HRM | = | high resolution melting |
| EOS | = | early onset sepsis |
| LOS | = | late onset sepsis |
| ELS | = | early onset bacterial sepsis followed by a late onset bacterial sepsis |
| GBS | = | Group B Streptococci |
| E.coli | = | Escherichia coli |
| CoNS | = | coagulase-negative Staphylococci |
| PCT | = | procalcitonin |
| GA | = | gestational age |
| NICU | = | neonatal intensive care unit |
| IMC | = | intermediate care unit |
| DBS | = | dried bloodspot filter card |
| TLR2 and TLR4 | = | Toll-like receptor 2 and 4 |
| MyD88 | = | Myeloid differentiation primary response |
| IL6 | = | interleukin-6 |
| IL6R | = | Interleukin-6 receptor |
| gp130 | = | Interleukin-6 signal transducer |
| IL8 | = | Interleukin-8 |
| CRP | = | C-reactive protein, pPROM, preterm premature rupture of membranes, BW, bodyweight |
| IGF2 | = | insulin-like growth factors 2 |
| Sp1 | = | specificity protein 1 |
Additional material
Download Zip (299.6 KB)Acknowledgments
The project was supported by the Oesterreichische Nationalbank (Anniversary Fund, project number: 14099). The present work benefited from the input of Mr S Pinezich, who provided valuable assistance in the undertaking of this project.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Supplemental Materials
Supplemental materials may be found here: http://www.landesbioscience.com/journals/epigenetics/article/26645/
References
- Cornell TT, Wynn J, Shanley TP, Wheeler DS, Wong HR. Mechanisms and regulation of the gene-expression response to sepsis. Pediatrics 2010; 125:1248 - 58; http://dx.doi.org/10.1542/peds.2009-3274; PMID: 20478944
- Edmond K, Zaidi A. New approaches to preventing, diagnosing, and treating neonatal sepsis. PLoS Med 2010; 7:e1000213; http://dx.doi.org/10.1371/journal.pmed.1000213; PMID: 20231868
- Bhutta ZA, Darmstadt GL, Hasan BS, Haws RA. Community-based interventions for improving perinatal and neonatal health outcomes in developing countries: a review of the evidence. Pediatrics 2005; 115:Suppl 519 - 617; PMID: 15866863
- Report WHO. 2012 - Levels and Trends in child mortality 2012.pdf. http://www.who.int/maternal_child_adolescent/documents/levels_trends_child_mortality_2012.pdf (accessed 16 Nov2012).
- Shane AL, Stoll BJ. Recent developments and current issues in the epidemiology, diagnosis, and management of bacterial and fungal neonatal sepsis. Am J Perinatol 2013; 30:131 - 41; http://dx.doi.org/10.1055/s-0032-1333413; PMID: 23297182
- Stoll BJ, Hansen N, Fanaroff AA, Wright LL, Carlo WA, Ehrenkranz RA, Lemons JA, Donovan EF, Stark AR, Tyson JE, et al. Late-onset sepsis in very low birth weight neonates: the experience of the NICHD Neonatal Research Network. Pediatrics 2002; 110:285 - 91; http://dx.doi.org/10.1542/peds.110.2.285; PMID: 12165580
- Curtis VA, Danquah LO, Aunger RV. Planned, motivated and habitual hygiene behaviour: an eleven country review. Health Educ Res 2009; 24:655 - 73; http://dx.doi.org/10.1093/her/cyp002; PMID: 19286894
- Weston EJ, Pondo T, Lewis MM, Martell-Cleary P, Morin C, Jewell B, Daily P, Apostol M, Petit S, Farley M, et al. The burden of invasive early-onset neonatal sepsis in the United States, 2005-2008. Pediatr Infect Dis J 2011; 30:937 - 41; http://dx.doi.org/10.1097/INF.0b013e318223bad2; PMID: 21654548
- Stoll BJ, Hansen NI, Sánchez PJ, Faix RG, Poindexter BB, Van Meurs KP, Bizzarro MJ, Goldberg RN, Frantz ID 3rd, Hale EC, et al, Eunice Kennedy Shriver National Institute of Child Health and Human Development Neonatal Research Network. Early onset neonatal sepsis: the burden of group B Streptococcal and E. coli disease continues. Pediatrics 2011; 127:817 - 26; http://dx.doi.org/10.1542/peds.2010-2217; PMID: 21518717
- Hornik CP, Fort P, Clark RH, Watt K, Benjamin DK Jr., Smith PB, Manzoni P, Jacqz-Aigrain E, Kaguelidou F, Cohen-Wolkowiez M. Early and late onset sepsis in very-low-birth-weight infants from a large group of neonatal intensive care units. Early Hum Dev 2012; 88:Suppl 2 S69 - 74; http://dx.doi.org/10.1016/S0378-3782(12)70019-1; PMID: 22633519
- Polin RA, Randis TM. Biomarkers for late-onset neonatal sepsis. Genome Med 2010; 2:58; http://dx.doi.org/10.1186/gm179; PMID: 20828428
- Kumar Y, Qunibi M, Neal TJ, Yoxall CW. Time to positivity of neonatal blood cultures. Arch Dis Child Fetal Neonatal Ed 2001; 85:F182 - 6; http://dx.doi.org/10.1136/fn.85.3.F182; PMID: 11668160
- Jörg Tost. DNA methylation: Methods and protocols. 2nd Ed.Humana Press: 2009. 3-20.
- Portela A, Esteller M. Epigenetic modifications and human disease. Nature Biotechnology. RE:view 2010; 28:1057 - 68
- Matwiyoff GN, Prahl JD, Miller RJ, Carmichael JJ, Amundson DE, Seda G, Daheshia M. Immune regulation of procalcitonin: a biomarker and mediator of infection. Inflamm Res 2012; 61:401 - 9; http://dx.doi.org/10.1007/s00011-012-0439-5; PMID: 22354317
- Schuetz P, Albrich W, Mueller B. Procalcitonin for diagnosis of infection and guide to antibiotic decisions: past, present and future. BMC Med 2011; 9:107; http://dx.doi.org/10.1186/1741-7015-9-107; PMID: 21936959
- Patil VK, Morjaria JB, De Villers F, Babu SK. Associations between procalcitonin and markers of bacterial sepsis. Medicina (Kaunas) 2012; 48:383 - 7; PMID: 23128457
- Oberhoffer M, Stonans I, Russwurm S, Stonane E, Vogelsang H, Junker U, Jäger L, Reinhart K. Procalcitonin expression in human peripheral blood mononuclear cells and its modulation by lipopolysaccharides and sepsis-related cytokines in vitro. J Lab Clin Med 1999; 134:49 - 55; http://dx.doi.org/10.1016/S0022-2143(99)90053-7; PMID: 10402059
- Sankar V, Webster NR. Clinical application of sepsis biomarkers. J Anesth 2013; 27:269 - 83; http://dx.doi.org/10.1007/s00540-012-1502-7; PMID: 23108494
- Bobetsis YA, Barros SP, Lin DM, Weidman JR, Dolinoy DC, Jirtle RL, Boggess KA, Beck JD, Offenbacher S. Bacterial infection promotes DNA hypermethylation. J. Dent Res 2007; 86:169 - 74; http://dx.doi.org/10.1177/154405910708600212
- Brodská H, Malíčková K, Adámková V, Benáková H, Šťastná MM, Zima T. Significantly higher procalcitonin levels could differentiate Gram-negative sepsis from Gram-positive and fungal sepsis. Clin Exp Med 2013; 13:165 - 70; http://dx.doi.org/10.1007/s10238-012-0191-8; PMID: 22644264
- Niwa T, Tsukamoto T, Toyoda T, Mori A, Tanaka H, Maekita T, Ichinose M, Tatematsu M, Ushijima T. Inflammatory processes triggered by Helicobacter pylori infection cause aberrant DNA methylation in gastric epithelial cells. Cancer Res 2010; 70:1430 - 40; http://dx.doi.org/10.1158/0008-5472.CAN-09-2755; PMID: 20124475
- Montagna MT, Coretti C, Rella A, Barbuti G, Manca F, Montagna O, Laforgia N, Caggiano G. The role of procalcitonin in neonatal intensive care unit patients with candidemia. Folia Microbiol (Praha) 2013; 58:27 - 31; http://dx.doi.org/10.1007/s12223-012-0169-7; PMID: 22688898
- Tan NY, Khachigian LM. Sp1 phosphorylation and its regulation of gene transcription. Mol Cell Biol 2009; 29:2483 - 8; http://dx.doi.org/10.1128/MCB.01828-08; PMID: 19273606
- Furuta T, Shuto T, Shimasaki S, Ohira Y, Suico MA, Gruenert DC, Kai H. DNA demethylation-dependent enhancement of toll-like receptor-2 gene expression in cystic fibrosis epithelial cells involves SP1-activated transcription. BMC Mol Biol 2008; 9:39; http://dx.doi.org/10.1186/1471-2199-9-39; PMID: 18423053
- Cooper DN, Mort M, Stenson PD, Ball EV, Chuzhanova NA. Methylation-mediated deamination of 5-methylcytosine appears to give rise to mutations causing human inherited disease in CpNpG trinucleotides, as well as in CpG dinucleotides. Hum Genomics 2010; 4:406 - 10; http://dx.doi.org/10.1186/1479-7364-4-6-406; PMID: 20846930
- Hur K, Niwa T, Toyoda T, Tsukamoto T, Tatematsu M, Yang HK, Ushijima T. Insufficient role of cell proliferation in aberrant DNA methylation induction and involvement of specific types of inflammation. Carcinogenesis 2011; 32:35 - 41; http://dx.doi.org/10.1093/carcin/bgq219; PMID: 20980348
- McClure EA, North CM, Kaminski NE, Goodman JI. Changes in DNA methylation and gene expression during 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced suppression of the lipopolysaccharide-stimulated IgM response in splenocytes. Toxicol Sci 2011; 120:339 - 48; http://dx.doi.org/10.1093/toxsci/kfq396; PMID: 21212295
- Kasper DC, Altiok I, Mechtler TP, Böhm J, Straub J, Langgartner M, Pollak A, Herkner KR, Berger A. Molecular detection of late-onset neonatal sepsis in premature infants using small blood volumes: proof-of-concept. Neonatology 2013; 103:268 - 73; http://dx.doi.org/10.1159/000346365; PMID: 23485823
- Li LC, Dahiya R. MethPrimer: designing primers for methylation PCRs. Bioinformatics 2002; 18:1427 - 31; http://dx.doi.org/10.1093/bioinformatics/18.11.1427; PMID: 12424112
- Wojdacz TK, Borgbo T, Hansen LL. Primer design versus PCR bias in methylation independent PCR amplifications. Epigenetics 2009; 4:231 - 4; PMID: 19483476