Abstract
Regulation of Set1-COMPASS-mediated H3K4 methylation and Dot1-mediated H3K79 methylation by H2BK123 ubiquitination (H2Bub1) is an evolutionarily conserved trans-histone crosstalk mechanism. How H2Bub1 impacts chromatin structure and affects Set1-COMPASS/Dot1 functions has not been fully defined. Ubiquitin was proposed to bind proteins to physically bridge H2Bub1 with Set1-COMPASS/Dot1. Alternatively, the bulky ubiquitin was thought to be a “wedge” that loosens the nucleosome for factor access. Contrary to the latter possibility, recent discoveries provide evidence for nucleosome stabilization by H2Bub1 via preventing the constant H2A-H2B eviction. Recent data has also uncovered a “docking-site” on H2B for Set1-COMPASS. Collectively, these findings invoke a model, where ubiquitin acts as a “glue” to bind the nucleosome together for supporting Set1-COMPASS/Dot1 functions. This review provides an overview of these novel findings. Additionally, how H2Bub1 and its deubiquitination might alter the chromatin dynamics during transcription is discussed. Possible models for nucleosome stabilization by ubiquitin are also provided.
Acknowledgements
We apologize for not citing other relevant articles due to space constraints. We thank Brian Strahl for his valuable suggestions. We thank The Vanderbilt-Ingram Cancer Center, The Robert J. and Helen C. Kleberg Foundation, NCI SPORE in Breast Cancer (5P0CA098131) and National Institutes of Health (RO1CA109355) for the financial support.
Figures and Tables
Figure 1 The trans-histone crosstalk between histone H2B ubiquitination and H3K4/-K79 methylation in budding yeast. A simulated structure for an ubiquitinated nucleosome is shown by placing the C-terminal residue of ubiquitin (PDB: 1UBQ) close to lysine 123 (K123) of H2B present in the yeast nucleosome (PDB 1ID3; H3, pale blue; H4, pale green; H2A, pale pink; H2B, pale yellow). Rad6 (the ubiquitin-conjugating enzyme) and Bre1 (the E3 ligase) conjugate ubiquitin onto K123 in the H2B C-terminal helix. The conjugated ubiquitin is removed by two deubiquitinases, Ubp8 and Ubp10. Histone methyltransferases Set1 and Dot1 catalyze mono- (me1), di- (me2) and trimethylation (me3) of H3 K4 and K79 residues, respectively. Set1 is present in a multi-protein complex (COMPASS) along with seven subunits, which regulate the complex stability, integrity and processive methylation.
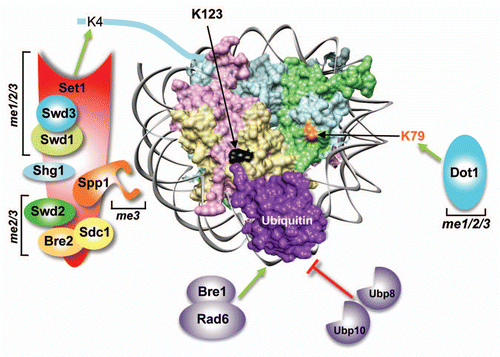
Figure 2 Model for the dynamic changes in yeast nucleosome stability mediated by H2B ubiquitination and deubiquitination. The innate weak intra-nucleosomal interactions and the constant H2A–H2B eviction lead to an unstable yeast nucleosome. Conjugation of ubiquitin stabilizes the nucleosome by negating these destabilizing events. On the contrary, removal of ubiquitin by deubiquitination promotes nucleosome instability. Stable nucleosome might allow or restrict factor binding to chromatin. By preventing H2A–H2B eviction, H2Bub1 retains the binding surfaces for Set1-COMPASS in H2A and H2B on chromatin. The composite “docking-site” for Set1-COMPASS on the nucleosome is predicted to be made up of the three residues in H2A (E65, N69 and D73) and the two experimentally confirmed residues in the H2B C-terminal helix (R119 and T122). Modified from Chandrasekharan et al.Citation59
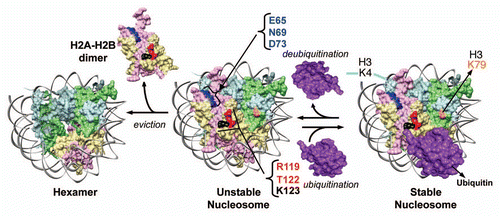
Figure 3 Model for the regulation of inducible gene expression by H2B ubiquitination and deubiquitination. (A) In budding yeast, conjugation of ubiquitin prevents H2A–H2B eviction and stabilizes the nucleosome positioned over the promoters of inducible genes in repressed state. The stable nucleosome might restrict the binding of activators (Act) and/or the TATA binding protein (TBP) to their cognate sites and prevents basal transcription. Following induction, DNA-bound activators recruit the SAGA complex containing the deubiquitinase Ubp8. Removal of the conjugated ubiquitin by Ubp8 causes nucleosome instability and promotes nucleosome disassembly by histone chaperones and/or other remodeling machineries. In turn, this allows the binding of TBP and recruitment of RNA Pol II for activated transcription. UAS, upstream activating sequence; TATA, the TATA box; INR, initiator sequence. (B) In mammals, inducible genes repressed by the E3 ligase RNF20 contain high levels of H2Bub1 and a poised RNA Pol II, and are enriched in the “marks” of active transcription [H3K9/14 acetylation (Ac, yellow circle) and H3K4me3 (cyan flag)]. H2Bub1 occupancy is high immediately downstream of the transcription start site and RNF20-repressed genes are in “closed”/compact chromatin configuration. Therefore, H2Bub1 mediated by RNF20 and possible E2-conjugating enzymes (UbcH6 or human homolog of Rad6, HR6A/B) might stabilize the nucleosome (PDB: 1KX5) and block transcription. USP22, the deubiquitinase in human SAGA (hSAGA), is required for the activation of many inducible genes. Therefore, following induction by activating stimuli, activators recruit hSAGA to the promoters of RNF-20 repressed genes to remove H2Bub1 and destabilize the nucleosome. This is followed by nucleosome disassembly, which results in “open”/loose chromatin formation. Removal of the stable “impeding” nucleosome allows the progression of paused RNA Pol II into the elongation phase. Adapted from Espinosa (2008).Citation46
![Figure 3 Model for the regulation of inducible gene expression by H2B ubiquitination and deubiquitination. (A) In budding yeast, conjugation of ubiquitin prevents H2A–H2B eviction and stabilizes the nucleosome positioned over the promoters of inducible genes in repressed state. The stable nucleosome might restrict the binding of activators (Act) and/or the TATA binding protein (TBP) to their cognate sites and prevents basal transcription. Following induction, DNA-bound activators recruit the SAGA complex containing the deubiquitinase Ubp8. Removal of the conjugated ubiquitin by Ubp8 causes nucleosome instability and promotes nucleosome disassembly by histone chaperones and/or other remodeling machineries. In turn, this allows the binding of TBP and recruitment of RNA Pol II for activated transcription. UAS, upstream activating sequence; TATA, the TATA box; INR, initiator sequence. (B) In mammals, inducible genes repressed by the E3 ligase RNF20 contain high levels of H2Bub1 and a poised RNA Pol II, and are enriched in the “marks” of active transcription [H3K9/14 acetylation (Ac, yellow circle) and H3K4me3 (cyan flag)]. H2Bub1 occupancy is high immediately downstream of the transcription start site and RNF20-repressed genes are in “closed”/compact chromatin configuration. Therefore, H2Bub1 mediated by RNF20 and possible E2-conjugating enzymes (UbcH6 or human homolog of Rad6, HR6A/B) might stabilize the nucleosome (PDB: 1KX5) and block transcription. USP22, the deubiquitinase in human SAGA (hSAGA), is required for the activation of many inducible genes. Therefore, following induction by activating stimuli, activators recruit hSAGA to the promoters of RNF-20 repressed genes to remove H2Bub1 and destabilize the nucleosome. This is followed by nucleosome disassembly, which results in “open”/loose chromatin formation. Removal of the stable “impeding” nucleosome allows the progression of paused RNA Pol II into the elongation phase. Adapted from Espinosa (2008).Citation46](/cms/asset/35f07e0e-71f0-4ae7-a988-012468d935c4/kepi_a_10912314_f0003.gif)
Figure 4 Model for the regulation of chromatin dynamics by H2B ubiquitination and deubiquitination during transcription elongation. (A) Rad6 and Bre1 associate with the Paf1 complex and travel with the elongating form of RNA Pol II, which is characterized by the phosphorylation (red oval) of serine residues in the heptad repeats that constitute the C-terminal domain (CTD, thick wavy line). (i and ii) Rad6/Bre1-mediated H2Bub1 stabilizes the nucleosome in front of the polymerase to counteract any torsional stress and might acts as a “checkpoint” to coordinate transcription-coupled events, such as, allowing the binding of Spt16/FACT and restricting Ctk1 binding to chromatin. (B) In (i), Ubp8, a component of the SAGA sub-complex that travels with RNA Pol II, removes the conjugated ubiquitin to destabilize the nucleosome. (ii) This facilitates nucleosome disassembly by Spt16/FACT. (iii) Deubiquitination by Ubp8 allows association of Ctk1 with chromatin for the phosphorylation of serine 2 in RNA Pol II CTD. (C) In (i), nucleosomes are reassembled behind the elongating RNA Pol II likely by the step-wise initial addition of H3-H4 by Spt6 followed by the addition of H2A–H2B by Spt16/FACT. (ii) Rad6/Bre1-mediated H2Bub1 stabilizes the nucleosome to facilitate nucleosome reassembly and to prevent any backtracking by RNA Pol II. (iii) Stable nucleosome also prevents any promiscuous transcription that might occur by the binding of TBP to cryptic TATA-like sequences within the coding region.
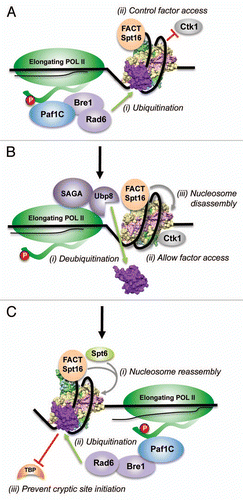
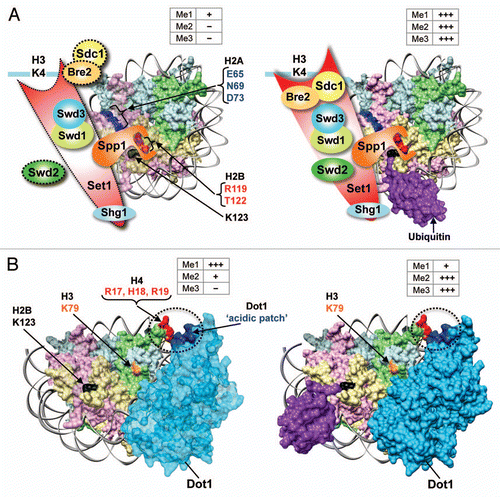
Figure 5 Model for the trans-histone crosstalk mechanism between H2Bub1 and H3K4/-K79 methylation. (A) Set1-COMPASS binds to the nucleosome via the interaction of Spp1 with the H2B C-terminal and likely by other hitherto unknown interactions with H2A. In the absence of H2Bub1, the constant H2A–H2B eviction and intrinsic nucleosome instability might disrupt or disassemble the composite “docking-site” for Set1-COMPASS on chromatin, leading to dissociation and/or weakened inter-subunit interactions within the complex and a catalytically-unfavorable conformation for Set1 (dotted lines). Together, these defects adversely affect the processive H3K4 methylation by Set1-COMPASS. In the presence of H2Bub1, H2A–H2B eviction is prevented and the stable nucleosome retains the binding surface for Set1-COMPASS on chromatin. This leads to increased chromatin association, inter-subunit interactions, and overall stability of Set1-COMPASS resulting in high levels of processive methylation. Modified from Chandrasekharan et al.Citation59 (B) Dot1 binds chromatin via the interaction of an acidic patch in its C-terminal region (EDVDE, amino acids 557–561) with a basic patch in H4 (RHR, amino acids 17–19). In the absence of H2Bub1, chromatin-bound Dot1 cannot catalyze H3K79me3 due to an inhibitory conformation (light blue). In the presence of H2Bub1, the stable nucleosome promotes an allosteric change in Dot1 that allows the production of high levels of H3K79me2 and -K79me3. +, low levels of methylation; +++, high levels of methylation; −, absence of methylation. Weak H2A–H2B dimer subjected to eviction, transparent and light colored; stable H2A–H2B dimer, bright colored.