
Abstract
Loss-of-function mutations in the methyl-DNA binding protein MeCP2 are associated with neurological dysfunction and impaired neural plasticity. However, the transcriptional changes that underlie these deficits remain poorly understood. Here, we show that mice bearing a C-terminal truncating mutation in Mecp2 (Mecp2308) are hypersensitive to the locomotor stimulating effects of cocaine. Furthermore, these mice have gene-specific alterations in striatal immediate-early gene (IEG) induction following cocaine administration. MeCP2 mutant mice show normal levels of baseline and cocaine-induced striatal Fos expression compared with their wild-type littermates. However, the mutant mice have enhanced cocaine-induced transcription of Junb and Arc. At the chromatin level, we find increased histone H3 acetylation at gene promoters in the Mecp2 mutant mice compared with their wild-type littermates, whereas two sites of repressive histone methylation are unchanged. Interestingly, we find that MeCP2 mutant mice show increased steady-state association of elongation-competent RNA Polymerase II (RNAP II) with the Junb and Arc promoters, whereas levels of RNAP II association at the Fos promoter are unchanged. These data reveal a gene-specific effect of MeCP2 on the recruitment of RNAP II to gene promoters that may modulate the inducibility of IEGs. In addition, our findings raise the possibility that aberrant regulation of IEGs including Junb and Arc may contribute to altered cocaine-induced neuronal and behavioral plasticity in Mecp2 mutant mice.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank A.N. Hutchinson, J.V. Deng and W.C. Wetsel for assistance with the animal experiments. This work was supported by NIH grant R01-DA022202 to A.E.W.
Figures and Tables
Figure 1 Mecp2 mutant mice are hypersensitive to the locomotor stimulating effects of cocaine. (A) Open field horizontal locomotor activity in Mecp2 WT and MUT mice. Mice were acclimated to the chamber then administered vehicle (saline) or 20 mg/kg cocaine (i.p.) after 60 min (black arrow). Horizontal locomotor activity was monitored for an additional 60 min. Lines show mean and error bars show SEM n = 9 WT vehicle, 5 MUT vehicle, 9 WT cocaine and 8 MUT cocaine. *p < 0.05 MUT cocaine vs. WT cocaine. (B) Quantification of cumulative locomotor activity in the 30 min following vehicle or cocaine administration. §p < 0.05 MUT vehicle vs. WT vehicle, *p < 0.05 WT cocaine vs. WT vehicle, #p < 0.05 MUT cocaine vs. WT cocaine, ^p < 0.05 MUT cocaine vs. MUT vehicle.
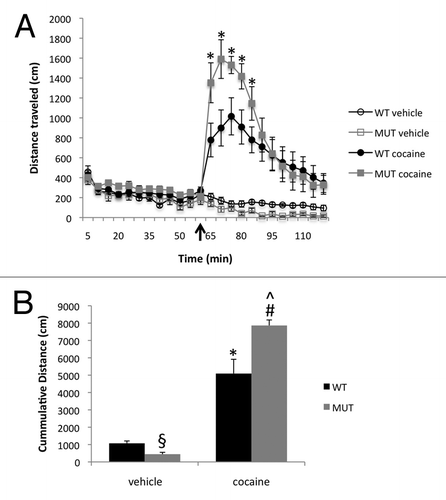
Figure 2 Enhanced cocaine-inducibility of Junb and Arc but not Fos following cocaine administration in Mecp2 mutant mice. Quantitative PC R was run on striatal lysates made from WT and MUT mice before or 1 h after administration of 20 mg/kg cocaine (i.p.). mRNA expression normalized to expression of the housekeeping gene Gapdh was scaled to basal levels in WT mice and presented as fold change. (A) Fos expression is induced by cocaine but does not differ between Mecp2 WT and MUT mice. (B) Junb expression is induced by cocaine and shows significantly greater induction by cocaine in Mecp2 MUT mice compared with WT. (C) Arc expression is induced by cocaine and shows significantly greater expression induction by cocaine in Mecp2 MUT mice compared with WT. (D) Tubb5 expression is not changed by cocaine administration and does not differ between WT and MUT mice. n = 6–8/group, *p < 0.05 WT cocaine vs. WT control, #p < 0.05 MUT cocaine vs. MUT control, ^p < 0.05 MUT cocaine vs. WT cocaine.
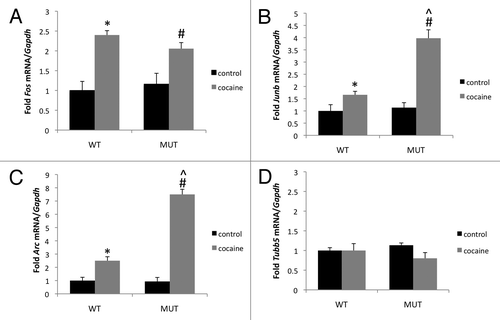
Figure 3 Enhanced histone H3 acetylation but no change in two sites of repressive histone H3 methylation at gene promoters in Mecp2 mutant mice. (A) ChIP was performed from striatum with an antibody that selectively recognizes histone H3 acetylated at lysine 9 and lysine 14 (H3K9/K14Ac). Pulldown quantities were normalized to the input for each experiment and relative quantities on each promoter were scaled to the constitutively transcribed gene Tubb5. (B) ChIP was performed from striatum with an antibody that selectively recognizes histone H3 trimethylated at lysine 27 (H3K27me3). Pulldown quantities were normalized to the input for each experiment and relative quantities on each promoter were scaled to the constitutively repressed gene Hbb-bh1. (C) ChIP was performed from striatum from an antibody that selectively recognizes histone H3 dimethylated at lysine 9 (H3K9me2). Pulldown quantities were normalized to the input for each experiment and relative quantities on each promoter were scaled to the constitutively repressed gene Hbb-bh1. n = 2 independent pools of 3–4 mice/group, *p < 0.05 MUT vs. WT.
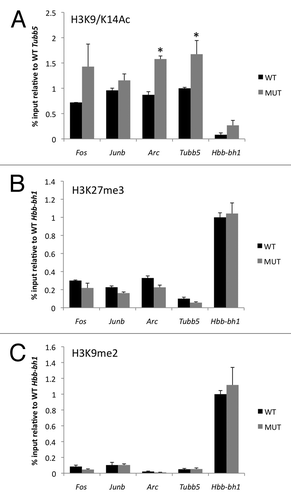
Figure 4 Enhanced association of elongation-competent RNA polymerase II with the Junb and Arc promoters in Mecp2 mutant mice. (A) ChIP was performed from striatum with an antibody that recognizes the large subunit of RNA Polymerase II. Pulldown quantities were normalized to the input for each experiment and relative quantities on each promoter were scaled to the constitutively expressed gene Tubb5. (B and C) ChIP was performed from striatum with antibodies that selectively recognize the Ser5 (B) or Ser2 (C) phosphorylated forms of the large subunit of RNA polymerase II. Pulldown quantities were normalized to the input for each experiment and relative quantities on each promoter are scaled to the constitutively expressed gene Tubb5. (D) No difference in pSer5:pSer2 ratio at IEG promoters in Mecp2 WT and MUT mice. (E) No difference in percentage of total PolII phosphorylated at Ser2 at IEG promoters in Mecp2 WT and MUT mice. n = 2 independent pools of 3–4 mice/group, *p < 0.05 MUT vs. WT.
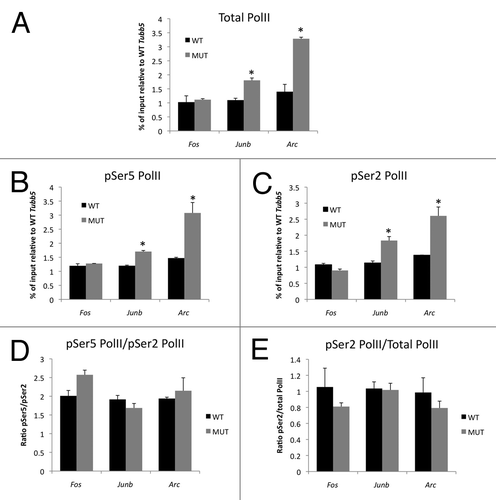
Table 1 Relative gene expression levels
Table 2 Gene expression primers used in this study
Table 3 ChIP primers used in this study