Abstract
Patients with inflammatory bowel diseases (IBD) harbour intestinal bacterial communities with altered composition compared with healthy counterparts; however, it is unknown whether changes in the microbiota are associated with genetic susceptibility of individuals for developing disease or instead reflect other changes in the intestinal environment related to the disease itself. Since deficiencies in the innate immune receptors Nod1 and Nod2 are linked to IBD, we tested the hypothesis that Nod-signaling alters intestinal immune profiles and subsequently alters bacterial community structure. We used qPCR to analyze expression patterns of selected immune mediators in the ileum and cecum of Nod-deficient mice compared with their Nod-sufficient littermates and assessed the relative abundance of major bacterial groups sampled from the ileum, cecum and colon. The Nod1-deficient ileum exhibited significantly lower expression of Nod2, Muc2, α- and β-defensins and keratinocyte-derived chemokine (KC), suggesting a weakened epithelial barrier compared with WT littermates; however, there were no significant differences in the relative abundance of targeted bacterial groups, indicating that Nod1-associated immune differences alone do not promote dysbiosis. Furthermore, Nod2-deficient mice did not display any changes in the expression of immune markers or bacterial communities. Shifts in bacterial communities that were observed in this study correlated with housing conditions and were independent of genotype. These findings emphasize the importance of using F2 littermate controls to minimize environmental sources of variation in microbial analyses, to establish baseline conditions for host-microbe homeostasis in Nod-deficient mice and to strengthen models for testing factors contributing to microbial dysbiosis associated with IBD.
Introduction
Intestinal homeostasis relies on a tightly regulated symbiosis between the host immune system and the resident microbiota. The importance of bacterial stimulation for immune maturation and normal physiological functioning is well documented in many studies using germ free mice.Citation1-Citation4 The innate immune receptors, Nod1 and Nod2, are the first line of defense for detection of bacterial peptidoglycan (PG) within the cytoplasm of epithelial cells. Upon PG binding, a common protein kinase, Rip2, is recruited, which in turn activates the NFκB signaling pathway leading to expression of genes including antimicrobial peptides, cytokines and chemokines, that contribute to strengthening the epithelial barrier. The Nod1 and Nod2 proteins are spatially and functionally distinct: whereas Nod1 appears to be expressed in all cells and recognizes the meso-diaminopimelate-d-alanine moiety of primarily Gram negative bacteria,Citation5,Citation6 Nod2 expression is thought to be in myeloid cells and Paneth cells and recognizes the muramyl dipeptide moiety found in the PG of both Gram positive and negative bacteria.Citation7,Citation8 These distinctions are not well understood with respect to their roles in regulating the intestinal microbiota. As a whole, the functions of Nod1 and Nod2 have been primarily demonstrated in the context of various inflammatory stimuli, including pathogenic microorganisms, where they have been shown to play key roles in regulating inflammation.Citation9 Their interactions with the resident intestinal bacteria and contributions to low levels of physiological inflammation remain poorly defined.
It follows that genetic defects that reduce or nullify functions of Nod1 or Nod2 proteins alter the immune profile of the intestinal ecosystem and potentially alter the composition and density of bacterial communities. Dysbiosis occurs when changes in bacterial community structure contribute to the development of disease, but the precise nature of these changes is not well understood. Indeed, genetic deficiencies in Nod1 and Nod2 are, respectively, linked to ulcerative colitis and Crohn disease, the two main manifestations of human IBD.Citation10,Citation11 These complex diseases are characterized by abnormal inflammatory responses and microbial dysbiosis, which may be initiated or enhanced by undefined environmental factors.Citation12,Citation13 Microbial dysbiosis in Crohn disease patients has been characterized by a higher abundance of bacteria from the Bacteroidetes and Proteobacteria phylaCitation14 and a lower abundance of bacteria from the Firmicutes phylum, particularly within the Clostridium XIVa and IV clusters,Citation15 compared with “healthy” counterparts. No spontaneous colitis occurs in Nod1- and Nod2-deficient mice housed in specific pathogen free (SPF) facilities. These conditions are ideal for testing hypotheses regarding the role of Nod-receptors in microbial homeostasis and dysbiosis.
Evidence from studies conducted in healthy SPF mice support a feedback model in which failure of Nod1 or Nod2 to activate the NFκB pathway for antimicrobial peptide secretion allows for expansion of certain bacterial groups. Using qPCR, Bouskra et al.Citation2 found that greater relative abundance of Bacteroides, Clostridales, SFB and Enterobacteriaceae (and lower abundance of Lactobacillaceae) groups correlated with reduced β-defensin (BD3) secretion in the terminal ileum of Nod1−/− mice compared with wild types (WT) housed in the same room. Similarly, Petnicki-Ocwieja et al.Citation16 reported greater abundance of Bacteroides and Firmicutes groups in the ileum (but not feces) of Nod2−/− compared with WT/Het mice from the same litter. This finding was linked to lower bactericidal capacity of crypt-secreted factors, such as α-defensins.Citation17 Abundance of Bacteroides was also greater in the ileum of Rip2−/− mice compared with WT littermates.Citation16 In contrast, Rehman et al.Citation18 reported greater abundance of Firmicutes in the ileum, and of Bacteroidetes in the feces, of Nod2−/− and WT mice derived, but separated at weaning, from the same heterozygous cross. From high-throughput sequencing analyses, Mondot et al.Citation19 found greater colonic abundance within the Rikenellaceae, Bacteroidaceae and Prevotellaceae families in Nod2−/− compared with WT mice with unspecified housing conditions. Recently, Couturier-Maillard et al.Citation20 reported an intrinsically dysbiotic microbiota in Nod2−/− and Rip2−/− mice that was transmissible to co-housed or co-fostered WT mice and increased their susceptibility to colitis and colitis-associated colorectal carcinoma following AOM and DSS treatment. Differences in methodology related to mouse breeding and housing, sampling intensity and location, and microbial analysis make it difficult to draw generalizable conclusions from these studies. Such methodological controls are imperative for establishing baseline conditions for subsequent experimentation, especially in light of recent findings that MyD88- and TLR- (TLR2, TLR4, TLR5, TLR9) deficient mouse colonies host distinct bacterial components in the small and large intestine that represent divergence of mouse strains after extended isolation but do not vary between littermates of different genotypes.Citation21
We tested the hypothesis that differences in intestinal immune profiles related to lack of Nod-mediated NFκB activation inherently alter the composition of gut bacterial communities at homeostasis. Moreover, we aimed to establish strict methodological controls for bacterial analysis and to explore potential sources of bacterial variation within our Nod1, Nod2 and Rip2 colonies. Results from this study will help establish baseline conditions on which to build future studies that examine the impact of innate signaling on the composition of the gut microbiota.
Results
Intestinal immune profiles associated with Nod deficiency at homeostasis
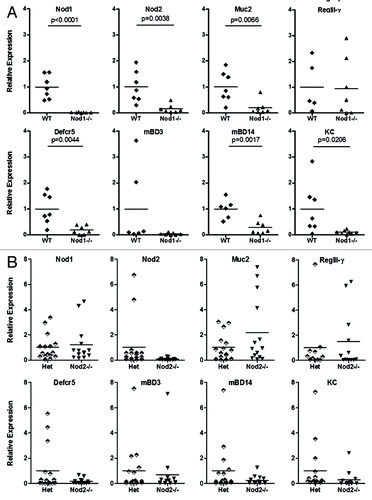
Using qPCR, we targeted a variety of genes () associated with innate immune functioning to establish baseline expression profiles in the Nod1−/− (n = 7) and Nod2−/− (n = 13) ileum and cecum relative to WT (n = 7) and Het (n = 16) littermates, respectively. The Nod1-deficient ileum was characterized by decreased relative expression of Nod2, mucin (Muc2), α- and β-defensins and KC, but no differences in RegIII-γ or cytokine (IL-6, IL-17A and IL-22) expression (). In contrast, no differences in any gene expression were found in the ileum of Nod2−/− compared with heterozygous littermates (). No altered expression profiles were found in cecum samples from either Nod1 or Nod2 mouse strains (data not shown). These findings suggest that Nod1 deficiency, but not Nod2, alters immune homeostasis within the ileal environment.
Table 1. Quantitative real time PCR primers for targeted mouse genes
Figure 1. Relative expression of target genes in the ileum of Nod1 (A) and Nod2 (B) littermates (mean control value set to 1).
No differences in bacterial abundance are related to Nod1 deficiency in C57BL/6 littermates
We next aimed to determine how lack of Nod1 and associated impacts on particular immune readouts, including Muc2 and Nod2 expression, affected microbial community composition. We used qPCR and 16S rDNA primers to target a range of bacterial groups () whose abundance has been correlated with altered immune functions or development of inflammatory disease.Citation22-Citation24
Table 2. Quantitative real time PCR primers for bacterial group-specific16S rDNA
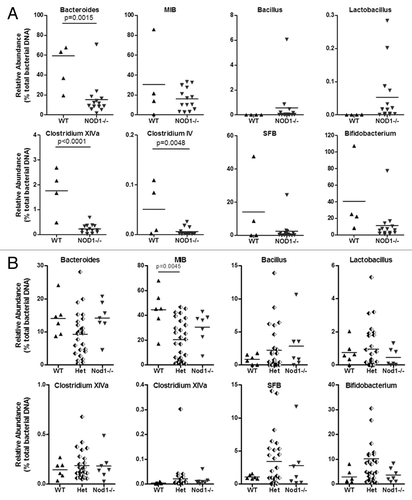
To determine the influence of mouse breeding and housing on the relative abundance of targeted bacterial groups, we analyzed 16S rDNA from WT (n = 5) and Nod1−/− (n = 16) mice housed in the same room (non-littermates), including eight co-caged females, as well as six litters of F2-generation mice consisting of WT (n = 6), Het (n = 27) and KO (n = 7) (littermates) genotypes. These analyses revealed significant differences in all three intestinal compartments, but only for the non-littermate group of mice. In the ileum, Nod1−/− mice had a lower abundance of Bacteroides, Clostridium XIVa and Clostridium IV groups and a trend toward higher Lactobacillus, compared with non-littermate control WT mice (). In the cecum, significantly lower abundance was found for only the MIB group, with trends toward lower abundance in both Clostridium groups (Fig. S1A). In the pellet, MIB and Bifidobacterium groups were both significantly lower and the Clostridium XIVa group also trended toward lower abundance (Fig. S2A). In contrast, no differences in bacterial abundance were found between Nod1−/− and WT littermates in any intestinal location ( and Fig. S1B and S2B). Enterobacteriaceae abundance was very low in all samples and did not vary between genotypes regardless of housing conditions (data not shown). There were no differences in bacterial density (total rRNA gene copies/g sample) calculated from the abundance of total Eubacteria DNA (data not shown). These data suggest that Nod1 deficiency and the altered immune environment in the intestine do not significantly impact on the relative abundance of targeted bacterial groups.
Figure 2. Relative abundance of targeted bacterial groups in the ileum of Nod1 non-littermates, including co-caged WT and Nod1−/− females (A) and Nod1 littermates (B).
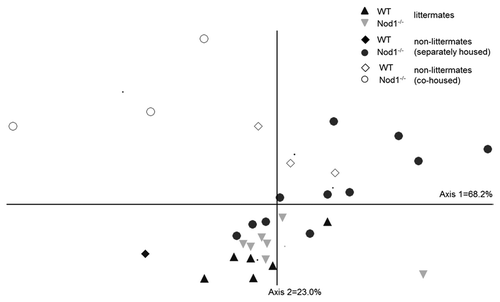
In order to further explore how familial transmission might impact on bacterial abundance between WT and Nod1-deficient mice, we performed nonmetric multidimensional scaling (NMS) analysis of the ileal bacterial communities. NMS showed tight clustering, indicating similarity, between the littermate Nod1−/− and WT mice (p = 0.316) and a more pronounced separation between the non-littermate genotypes (). Three weeks of co-caging WT and Nod1−/− non-littermates shifted the community structure of the Nod1−/− mice away from the mean community structure of the separated mice (p = 0.0018) and this trend is also seen for WT mice. Bacterial communities from co-caged, non-littermate Nod1−/− and WT mice exhibited high variation and were significantly different from each other (p = 0.0252). In the cecum and colon, differences in community structure were not sufficient to spatially separate mice into genotype or housing-related groups. This analysis further strengthens the notion that familial relatedness, more than Nod1-deficiency, shapes the bacterial community structure within the intestine.
Figure 3. NMS ordination of bacterial community structure in the ileum of WT and Nod1−/− mice under different housing conditions. Littermates tend to cluster closely together while there is greater separation between genotypes of the non-littermate mice. Co-caged mice (open symbols) tend to separate from the other groups.
Bacterial abundance profiles do not vary between littermates from Nod2- and Rip2-deficient mouse strains
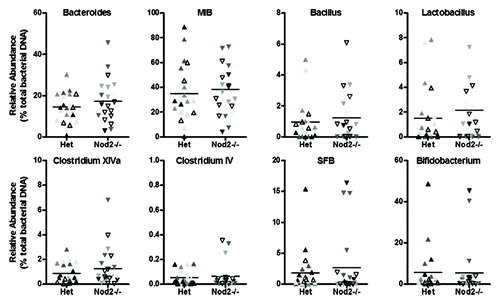
We next assessed whether bacterial abundance varied in our Nod2 and Rip2 strains. For Nod2-deficient mice, we compared relative abundance in Nod2−/− (n = 22) and heterozygous (n = 23) littermates (five litters) since a previous study reported no differences between WT and heterozygous littermates.Citation16 No differences in relative abundance of any bacterial group were found between genotypes in the ileum (), cecum or pellet (Fig. S3 and 4) samples and variation between individual mice was consistent between litters.
Figure 4. Relative abundance of targeted bacterial groups in the ileum of Het and Nod2−/− littermates. Variation within litters is shown by the different shading of points on each graph.
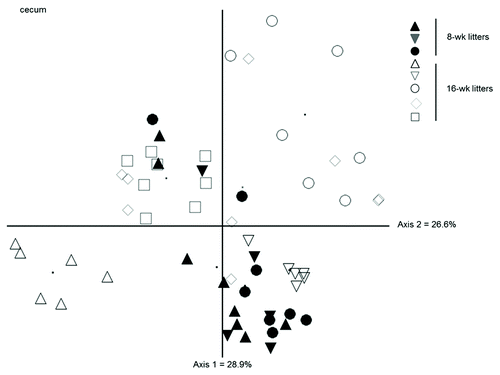
For the Rip2 strain, we analyzed three litters of 8-week-old mice and five litters of 16-week-old mice and found no differences in relative bacterial abundance in any intestinal compartment or at either age (data not shown). As we used a more restricted primer set for the 8-week samples, we could not directly compare community structure between age groups. Instead, an approximately 300 bp fragment of 16S rDNA, including variable regions V1 and V2, was amplified from all samples using general bacterial primers and analyzed by length heterogeneity PCR (LH-PCR), a fingerprinting method that separates the bacterial community based on inherent length differences in the 16S rRNA gene.Citation25 The NMS ordination () shows the effect of litter on the community structure of cecum samples, regardless of genotypes. Whereas litters 1−3 (8-week mice) do not vary significantly from one another, each 16-week litter varies from every other 16-week litter as well as from all of the 8-wk litters (p < 0.05). These findings are consistent with the idea that neither lack of Nod2 or Rip2 significantly impacts on the intestinal bacterial communities in controlled littermate analyses and that extended separation of mice tends to augment variation between and within litters.
Figure 5. NMS ordination of bacterial community structure (from fragment analysis by LH-PCR) in the cecum of WT and Rip2−/− showing variation between litters at 8 and 16 weeks. The community structure of 8-week mice did not vary significantly from each other but the 16-week mice varied from the 8-week mice as well as with each other, regardless of genotype.
Discussion
Our study highlights the inherent stability of bacterial communities inhabiting the ileum, cecum and colon of Nod1−/−, Nod2−/− and Rip2−/− and their WT or heterozygous littermates under homeostatic conditions. For the bacterial groups that we targeted, this stability persisted despite the presence of an altered immune environment in the Nod1-deficient ileum, which could contribute to a weakened epithelial barrier. Stability in community structure was also reported from DGGE fingerprinting of colon samples from Nod1Nod2 DKO and Het-Het littermates despite decreased expression of antimicrobial RegIII-γ and E-cadherin and increased paracellular permeability, other markers of a weakened epithelial barrier.Citation26 Our results are also in line with a recent report that found no differences in bacterial community structure between MyD88-, TLR2-, TLR4-, TLR5- and TLR9-deficient and their WT littermates using high-throughput sequencing.Citation21 These findings emphasize the need for robust experimental design for evaluating intestinal bacterial communities. In particular, performing analysis on F2 littermates of heterozygous crosses is imperative before making claims that a particular genotype influences the composition of the gut microbiota. Thus, our results suggest that altered bacterial abundance patterns reported in other studies on Nod-deficient mice might be a reflection of methodological differences and lack of proper controls.
As previously mentioned, of primary importance for study design is to include use of F2 littermate controls and adequate numbers of mice to compensate for high variation. In our comparison of non-littermate, including co-caged, and littermate sets of mice, we found significant differences in relative abundance (decreased Bacteroides, Clostidium groups XIVa and IV (ileum), decreased MIB (cecum and pellet) and decreased Bifidobacterium (pellet)) in the non-littermates that were not present in the littermate analysis. From this, and also from analyses of Nod2 and Rip2 littermates that also showed no abundance differences, it is clear that the composition of the intestinal microbiota is determined by the long-term, shared environment of littermate mice.Citation27 The community structure (NMS) analysis, based on abundance of the nine targeted bacterial groups, confirmed the lack of differences between WT and KO littermates and showed a more pronounced separation of the non-littermate set of mice. Furthermore, co-caging of non-littermate WT and KO females resulted in community structures that were variable and distinct from other mice of the same genotype. This variation may reflect a dynamic re-adjustment of microbial community structure in the co-caged mice to adapt to novel bacterial species/ strains associated with the isolated mouse strains into each other’s established intestinal communities.
Anatomic location is another of the dominant sources of variation in bacterial community structure and variations within distinct intestinal habitats are not well understood at different spatial scales.Citation28 Bacterial abundance varied across the three intestinal locations that we sampled (mucosa of the distal ileum, cecum contents and fecal pellet), but no sets of Nod1, Nod2 or Rip2 littermates exhibited any significant abundance shifts in any intestinal compartment. It is possible that our sampling methods were insufficiently precise or introduced too much noise to detect differences in bacterial abundance related to lack of Nod2 activation, as these effects might be spatially confined to intestinal crypts or deeper intestinal layers.Citation16,Citation17 However, since sampling location/ method was similar to sampling units in other studies,Citation16,Citation18,Citation19 it is likely that discrepancies in bacterial abundance assessments are related to other differences in methodology.
We also used qPCR to characterize the immune environment of Nod1- and Nod2-deficient mice. While Nod2-deficient mice did not present any differences in gene expression compared with heterozygous littermates, the immune profile of the Nod1-deficient ileum suggested a more susceptible epithelial barrier, characterized by decreased expression of both Nod1 and Nod2, mucin, α- and β-defensins and KC. These effects are subtle, reflecting only 2–3 fold differences, but taken together, may represent an important vulnerability of the Nod1 ileal environment to external perturbation. Although a disease of the colon, ulcerative colitis is confined to the mucosal layers and has been specifically associated with epithelial barrier genes in contrast to Crohn’s disease, which impacts the deeper layers of the gut wall.Citation29 Lower expression of β-defensins has been previously associated with Nod1 deficiency and linked to greater density of bacteria at the mucosal surface.Citation2,Citation30 However, lower expression of the α-defensin, Defcr5, an antimicrobial peptide preferentially produced in intestinal Paneth cells but expressed normally in Nod2−/− miceCitation17 has not been previously related to Nod1 function. Mucin (Muc2) is a major component of mucus that covers the small intestine, which, unlike the mucus of the colon, is not attached to the epithelium and is permeable to bacteria.Citation31 Decreased secretion of the mucus may allow bacteria access to the epithelium. The lower expression of Nod2 in Nod1-deficient mice was unexpected since we hypothesized that, because of the partial overlap in their sensing specificities, there might be compensatory induction of Nod2. Indeed, the diminished expression of Nod2 in the Nod1-deficient intestinal environment might contribute to the lower expression of protective factors compared with WT animals.
Little is known about how the host’s immune system regulates interactions with the intestinal microbiota. Studies from Crohn disease patients support the idea that an altered immune environment within the intestine fosters dysbiosis. However, the altered microbiota in disease is likely shaped by the chronic inflammatory state of the intestine, as a result of both genetic and environmental factors. Indeed, whether dysbiosis is present before disease onset is unknown. Our results suggest that at least in baseline conditions, lack of Nod-mediated signals do not significantly impact on bacterial communities in the gut. Instead, other components of the innate and adaptive immune system, acting separately or in synergy, may play a larger role in controlling the microbiota and maintaining a balanced environment.Citation32 For example, MyD88-dependent pathways that control the expression of several other antimicrobial factors, including RegIII-γ, RegIII-β, RELMβ, CRP-ductin, are likely more important for controlling bacterial penetration of the intestinal barrier and maintaining homeostasis.Citation33,Citation34
Our findings support the notion that expression of Nod1, Nod2 or Rip2 does not significantly impact on the structure or abundance of intestinal bacterial communities at homeostasis when all other environmental variables are controlled. We highlight the importance for using robust methodology to establish the baseline conditions in mouse strains of interest, as only then will it be possible to experimentally test the effects of exogenous factors on the limits of community stability (resistance to change) and resilience (ability to recover following disturbance) in immune-deficient mouse models. Ecological insights into the intestinal microbiome will lead to a greater understanding of the links with chronic inflammatory disease but meaningful insights will only be gained if appropriate experimental designs are applied.
Methods
Mice
C57BL/6 mice were housed in the specific pathogen free (SPF) mouse facility at the University of Toronto. Wild type (WT) mice were crossed with Nod1−/−, Nod2−/− or Rip2−/− knockout (KO) mice to generate F1 heterozygotes for each KO mouse strain. Other WT (n = 5) and Nod1−/− (n = 24) mice from our colony were maintained separately and exposed to fecal material from all other cages. Of these, eight females (n = 4 for WT and KO) were co-caged for three weeks prior to sampling and analysis of the entire group. For the Nod1 and Rip2 mouse strains, three breeding pairs of F1 heterozygotes were established to generate F2 littermates comprised of WT, Het and KO genotypes. For the Nod2 strain, two pairs of F1 heterozygotes were crossed with KO to produce F2 littermates comprised of Het and KO genotypes. All mice were euthanized at 8–10 weeks (and 16 weeks in the case of Rip2) and multiple litters from each set of breeding pairs were sampled. Sample sizes for control and KO mice ranged from 3–27 for all analyses.
Samples from each mouse included mucosal scrapings from a 12-cm length of the terminal ileum, cecum contents and the fecal pellet (from the lower colon/ rectum) for bacterial analysis and tissue from the ileum and cecum to assess host gene expression.
Quantitative real time PCR to analyze relative gene expression
RNA was extracted from ileum and cecum tissues using the RNeasy Mini Kit (Qiagen cat. 74104), treated with TURBOTM DNase (Life Technologies cat. AM2238) and converted to cDNA using the SuperScript® III First-Strand Synthesis SuperMix for qRT-PCR (Life Technologies cat. 11752050) following manufacturer instructions. The cDNA (~10 ng/μl) was analyzed by qPCR using primer sets targeting several mouse genes related to proper functioning of the epithelial barrier (). These include genes encoding: the Nod1 and Nod2 receptor proteins; the Muc2 protein constituent of mucus lining the intestinal epitheliaCitation31; RegIII-γ, a bactericidal C-type lectin that specifically targets Gram-positive bacteriaCitation34; antimicrobial peptides, including the α-defensin (cryptin) Defcr5 and the β-defensins BD3 and BD14; the chemokine CXCL1 (KC), which is a chemotactic factor involved in neutrophil recruitmentCitation35; and several pro-inflammatory cytokines, IL-6, IL-17a and IL-22. Quantitative PCR was performed using Power SYBR® Green PCR Master Mix (Life Technologies cat. 4367659) in a ViiA™ 7 Real-Time PCR System (Applied Biosystems) under the following run conditions: initial denaturation at 95°C for 10 min, followed by 40 cycles of 15 sec at 95°C and 1 min at 60°C. Data were acquired in the final step at 60°C and melting curves (65 to 95°C) were generated for each set of primers. Relative gene expression was calculated using the ΔCt method and normalizing to the rpl19 housekeeping gene. For comparison of gene expression between WT and KO mice, data was expressed as a fold change relative to the WT condition which was set at 1.
Quantitative real time PCR to analyze relative abundance of bacterial groups
Total DNA was extracted from the ileum, cecum and pellet samples using the MoBio PowerSoil kit (MoBio, cat. 12888) and following manufacturer’s instructions. Bacterial DNA (~10 ng/μl) was analyzed by qPCR as described in the previous section using 16S rDNA primers (Integrated DNA Technologies) to target specific groups (). These include members of the Gram-negative Bacteroidetes (Bacteroides and Mouse Intestinal Bacteroides (MIB) groups) and Proteobacteria (Enterobacteriaceae group) phyla and members of the Gram-positive Firmicutes (Bacillus, Lactobacillus, Clostridium cluster XIVa, Clostridium cluster IV and segmented filamentous bacteria (SFB) groups) and Actinobacteria (Bifidobacterium group) phyla. Relative abundance of bacterial groups was calculated by normalizing ΔCt for each target group to the Eubacteria (housekeeper) group. Bacterial density was calculated from the total number of 16S rRNA gene copies (Eubacteria) based on each sample’s DNA concentration and weight.Citation36 However, high error (e.g., the variability of 16S rRNA gene copy number across taxa, procedural biases, etc.) associated with this density calculation make it less sensitive for detecting small differences.
Length heterogeneity PCR
DNA from the cecum samples of 8-and 16-wk Rip2 littermates was amplified by PCR using the FAM-labeled forward primer 27F (5′-AGAGTTTGATCMTGGCTCAG) and unlabeled reverse primer 338R (5′-GCTGCCTCCCGTAGGAGT).Citation25 Reactions included preliminary denaturation for 3 min at 94°C, followed by 30 cycles of denaturation at 94°C for 1 min, annealing 53°C for 1 min and elongation at 72°C for 1 min and a final elongation step at 72°C for 10 min. PCR products were cleaned by ethanol precipitation and then loaded into a ABI PRISM® 3730XL Genetic Analyzer along with 400 GeneScanT 400 LIZ® (Applied Biosystems) size standard mixture. Raw data was analyzed using GeneScan® Analysis v3.5.1 (Applied Biosystems). Operational taxonomic units (OTUs) were determined after applying a threshold of 1% to the total intensity of the sample and assigning the remaining OTUs to bins of 3 bp (± 1 bp).
Statistics
Relative bacterial abundance and host gene expression were compared between WT, Het and KO groups using either T-tests or ANOVA (α = 0.05), depending on the number of groups. Bacterial community structure (at the level of the mouse intestinal compartment) was assessed graphically with Nonmetric Multidimensional Scaling (NMS) using PC-ORD 6.0 software.Citation37 NMS was calculated from abundance matrices for the nine targeted groups (qPCR data) and OTUs (LH-PCR data) on the basis of a Sørensen (Bray-Curtis) distance measure with 250 runs with real and randomized data (compared using Monte Carlo simulations) and a maximum of 500 iterations to assess stability.Citation38 A stepwise reduction in dimensionality was used to minimize stress along with a random starting configuration. Pairwise comparisons between groups were tested with Multi-Response Permutation Procedures (MRPP) using the Sørensen distance measure.
| Abbreviations: | ||
| AOM | = | azoxymethane |
| DSS | = | dextran sodium sulfate |
| IBD | = | inflammatory bowel disease |
| KO | = | knockout |
| LH-PCR | = | length heterogeneity polymerase chain reaction |
| OTU | = | operational taxonomic unit |
| MRPP | = | Multi-Response Permutation Procedures |
| NMS | = | Nonmetric Multidimensional Scaling |
| PG | = | peptidoglycan |
| SPF | = | specific pathogen free |
| WT | = | wild type |
Additional material
Download Zip (615.2 KB)Acknowledgments
This work was supported by grants from the Canadian Institutes of Health Research, Crohn's and Colitis Foundation of Canada and Canadian Association of Gastoenterology. We thank H. Mascarenhas for assistance with sample processing and Dr H. Maughan for valuable discussions.
Disclosure of Potential Conflicts of Interest
No potential conflict of interest was disclosed.
References
- Mazmanian SK, Liu CH, Tzianabos AO, Kasper DL. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005; 122:107 - 18; http://dx.doi.org/10.1016/j.cell.2005.05.007; PMID: 16009137
- Bouskra D, Brézillon C, Bérard M, Werts C, Varona R, Boneca IG, et al. Lymphoid tissue genesis induced by commensals through NOD1 regulates intestinal homeostasis. Nature 2008; 456:507 - 10; http://dx.doi.org/10.1038/nature07450; PMID: 18987631
- Gaboriau-Routhiau V, Rakotobe S, Lécuyer E, Mulder I, Lan A, Bridonneau C, et al. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 2009; 31:677 - 89; http://dx.doi.org/10.1016/j.immuni.2009.08.020; PMID: 19833089
- Ganal SC, Sanos SL, Kallfass C, Oberle K, Johner C, Kirschning C, et al. Priming of natural killer cells by nonmucosal mononuclear phagocytes requires instructive signals from commensal microbiota. Immunity 2012; 37:171 - 86; http://dx.doi.org/10.1016/j.immuni.2012.05.020; PMID: 22749822
- Chamaillard M, Hashimoto M, Horie Y, Masumoto J, Qiu S, Saab L, et al. An essential role for NOD1 in host recognition of bacterial peptidoglycan containing diaminopimelic acid. Nat Immunol 2003; 4:702 - 7; http://dx.doi.org/10.1038/ni945; PMID: 12796777
- Girardin SE, Boneca IG, Carneiro LA, Antignac A, Jéhanno M, Viala J, et al. Nod1 detects a unique muropeptide from gram-negative bacterial peptidoglycan. Science 2003; 300:1584 - 7; http://dx.doi.org/10.1126/science.1084677; PMID: 12791997
- Girardin SE, Boneca IG, Viala J, Chamaillard M, Labigne A, Thomas G, et al. Nod2 is a general sensor of peptidoglycan through muramyl dipeptide (MDP) detection. J Biol Chem 2003; 278:8869 - 72; http://dx.doi.org/10.1074/jbc.C200651200; PMID: 12527755
- Inohara N, Ogura Y, Fontalba A, Gutierrez O, Pons F, Crespo J, et al. Host recognition of bacterial muramyl dipeptide mediated through NOD2. Implications for Crohn’s disease. J Biol Chem 2003; 278:5509 - 12; http://dx.doi.org/10.1074/jbc.C200673200; PMID: 12514169
- Rubino SJ, Selvanantham T, Girardin SE, Philpott DJ. Nod-like receptors in the control of intestinal inflammation. Curr Opin Immunol 2012; 24:398 - 404; http://dx.doi.org/10.1016/j.coi.2012.04.010; PMID: 22677577
- Hugot JP, Chamaillard M, Zouali H, Lesage S, Cézard JP, Belaiche J, et al. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn’s disease. Nature 2001; 411:599 - 603; http://dx.doi.org/10.1038/35079107; PMID: 11385576
- McGovern DP, Hysi P, Ahmad T, van Heel DA, Moffatt MF, Carey A, et al. Association between a complex insertion/deletion polymorphism in NOD1 (CARD4) and susceptibility to inflammatory bowel disease. Hum Mol Genet 2005; 14:1245 - 50; http://dx.doi.org/10.1093/hmg/ddi135; PMID: 15790594
- Sartor RB. Microbial influences in inflammatory bowel diseases. Gastroenterology 2008; 134:577 - 94; http://dx.doi.org/10.1053/j.gastro.2007.11.059; PMID: 18242222
- Maloy KJ, Powrie F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011; 474:298 - 306; http://dx.doi.org/10.1038/nature10208; PMID: 21677746
- Eckburg PB, Relman DA. The role of microbes in Crohn’s disease. Clin Infect Dis 2007; 44:256 - 62; http://dx.doi.org/10.1086/510385; PMID: 17173227
- Frank DN, Robertson CE, Hamm CM, Kpadeh Z, Zhang T, Chen H, et al. Disease phenotype and genotype are associated with shifts in intestinal-associated microbiota in inflammatory bowel diseases. Inflamm Bowel Dis 2011; 17:179 - 84; http://dx.doi.org/10.1002/ibd.21339; PMID: 20839241
- Petnicki-Ocwieja T, Hrncir T, Liu YJ, Biswas A, Hudcovic T, Tlaskalova-Hogenova H, et al. Nod2 is required for the regulation of commensal microbiota in the intestine. Proc Natl Acad Sci U S A 2009; 106:15813 - 8; http://dx.doi.org/10.1073/pnas.0907722106; PMID: 19805227
- Kobayashi KS, Chamaillard M, Ogura Y, Henegariu O, Inohara N, Nuñez G, et al. Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science 2005; 307:731 - 4; http://dx.doi.org/10.1126/science.1104911; PMID: 15692051
- Rehman A, Sina C, Gavrilova O, Häsler R, Ott S, Baines JF, et al. Nod2 is essential for temporal development of intestinal microbial communities. Gut 2011; 60:1354 - 62; http://dx.doi.org/10.1136/gut.2010.216259; PMID: 21421666
- Mondot S, Barreau F, Al Nabhani Z, Dussaillant M, Le Roux K, Doré J, et al. Altered gut microbiota composition in immune-impaired Nod2(-/-) mice. Gut 2012; 61:634 - 5; http://dx.doi.org/10.1136/gutjnl-2011-300478; PMID: 21868489
- Couturier-Maillard A, Secher T, Rehman A, Normand S, De Arcangelis A, Haesler R, et al. NOD2-mediated dysbiosis predisposes mice to transmissible colitis and colorectal cancer. J Clin Inv 2013.
- Ubeda C, Lipuma L, Gobourne A, Viale A, Leiner I, Equinda M, et al. Familial transmission rather than defective innate immunity shapes the distinct intestinal microbiota of TLR-deficient mice. J Exp Med 2012; 209:1445 - 56; http://dx.doi.org/10.1084/jem.20120504; PMID: 22826298
- Bloom SM, Bijanki VN, Nava GM, Sun L, Malvin NP, Donermeyer DL, et al. Commensal Bacteroides species induce colitis in host-genotype-specific fashion in a mouse model of inflammatory bowel disease. Cell Host Microbe 2011; 9:390 - 403; http://dx.doi.org/10.1016/j.chom.2011.04.009; PMID: 21575910
- Dongarra ML, Rizzello V, Muccio L, Fries W, Cascio A, Bonaccorsi I, et al. Mucosal Immunology and Probiotics. Curr Allergy Asthma Rep 2012; 13:19 - 26; http://dx.doi.org/10.1007/s11882-012-0313-0; PMID: 23054627
- Hooper LV, Littman DR, Macpherson AJ. Interactions between the microbiota and the immune system. Science 2012; 336:1268 - 73; http://dx.doi.org/10.1126/science.1223490; PMID: 22674334
- Kovacs A, Ben-Jacob N, Tayem H, Halperin E, Iraqi FA, Gophna U. Genotype is a stronger determinant than sex of the mouse gut microbiota. Microb Ecol 2011; 61:423 - 8; http://dx.doi.org/10.1007/s00248-010-9787-2; PMID: 21181142
- Natividad JM, Petit V, Huang X, de Palma G, Jury J, Sanz Y, et al. Commensal and probiotic bacteria influence intestinal barrier function and susceptibility to colitis in Nod1-/-; Nod2-/- mice. Inflamm Bowel Dis 2012; 18:1434 - 46; http://dx.doi.org/10.1002/ibd.22848; PMID: 22162005
- Costello EK, Stagaman K, Dethlefsen L, Bohannan BJ, Relman DA. The application of ecological theory toward an understanding of the human microbiome. Science 2012; 336:1255 - 62; http://dx.doi.org/10.1126/science.1224203; PMID: 22674335
- Relman DA. The human microbiome: ecosystem resilience and health. Nutr Rev 2012; 70:Suppl 1 S2 - 9; http://dx.doi.org/10.1111/j.1753-4887.2012.00489.x; PMID: 22861804
- Parkes M. Evidence from genetics for a role of autophagy and innate immunity in IBD pathogenesis. Dig Dis 2012; 30:330 - 3; http://dx.doi.org/10.1159/000338119; PMID: 22796792
- Grubman A, Kaparakis M, Viala J, Allison C, Badea L, Karrar A, et al. The innate immune molecule, NOD1, regulates direct killing of Helicobacter pylori by antimicrobial peptides. Cell Microbiol 2010; 12:626 - 39; http://dx.doi.org/10.1111/j.1462-5822.2009.01421.x; PMID: 20039881
- Johansson ME, Hansson GC. Microbiology. Keeping bacteria at a distance. Science 2011; 334:182 - 3; http://dx.doi.org/10.1126/science.1213909; PMID: 21998374
- Hansen J, Gulati A, Sartor RB. The role of mucosal immunity and host genetics in defining intestinal commensal bacteria. Curr Opin Gastroenterol 2010; 26:564 - 71; http://dx.doi.org/10.1097/MOG.0b013e32833f1195; PMID: 20871399
- Vaishnava S, Behrendt CL, Ismail AS, Eckmann L, Hooper LV. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interface. Proc Natl Acad Sci U S A 2008; 105:20858 - 63; http://dx.doi.org/10.1073/pnas.0808723105; PMID: 19075245
- Vaishnava S, Yamamoto M, Severson KM, Ruhn KA, Yu X, Koren O, et al. The antibacterial lectin RegIIIgamma promotes the spatial segregation of microbiota and host in the intestine. Science 2011; 334:255 - 8; http://dx.doi.org/10.1126/science.1209791; PMID: 21998396
- Loving CL, Osorio M, Kim YG, Nuñez G, Hughes MA, Merkel TJ. Nod1/Nod2-mediated recognition plays a critical role in induction of adaptive immunity to anthrax after aerosol exposure. Infect Immun 2009; 77:4529 - 37; http://dx.doi.org/10.1128/IAI.00563-09; PMID: 19620350
- Wlodarska M, Willing B, Keeney KM, Menendez A, Bergstrom KS, Gill N, et al. Antibiotic treatment alters the colonic mucus layer and predisposes the host to exacerbated Citrobacter rodentium-induced colitis. Infect Immun 2011; 79:1536 - 45; http://dx.doi.org/10.1128/IAI.01104-10; PMID: 21321077
- McCune B, Mefford MJPC-ORD. Multivariate Analysis of Ecological Data. MjM Software Design, 1999.
- McCune B, Grace JB, Urban DL. Analysis of Ecological Communities. MjM Software Design Gleneden Beach, Oregon, 2002.
- Zheng Y, Valdez PA, Danilenko DM, Hu Y, Sa SM, Gong Q, et al. Interleukin-22 mediates early host defense against attaching and effacing bacterial pathogens. Nat Med 2008; 14:282 - 9; http://dx.doi.org/10.1038/nm1720; PMID: 18264109
- Duary RK, Bhausaheb MA, Batish VK, Grover S. Anti-inflammatory and immunomodulatory efficacy of indigenous probiotic Lactobacillus plantarum Lp91 in colitis mouse model. Mol Biol Rep 2012; 39:4765 - 75; http://dx.doi.org/10.1007/s11033-011-1269-1; PMID: 21947851
- Tebar LA, Géranton SM, Parsons-Perez C, Fisher AS, Bayne R, Smith AJ, et al. Deletion of the mouse RegIIIbeta (Reg2) gene disrupts ciliary neurotrophic factor signaling and delays myelination of mouse cranial motor neurons. Proc Natl Acad Sci U S A 2008; 105:11400 - 5; http://dx.doi.org/10.1073/pnas.0711978105; PMID: 18678917
- Steinbrecher KA, Harmel-Laws E, Sitcheran R, Baldwin AS. Loss of epithelial RelA results in deregulated intestinal proliferative/apoptotic homeostasis and susceptibility to inflammation. J Immunol 2008; 180:2588 - 99; PMID: 18250470
- Burd RS, Furrer JL, Sullivan J, Smith AL. Murine beta-defensin-3 is an inducible peptide with limited tissue expression and broad-spectrum antimicrobial activity. Shock 2002; 18:461 - 4; http://dx.doi.org/10.1097/00024382-200211000-00013; PMID: 12412627
- Sørensen OE, Thapa DR, Roupé KM, Valore EV, Sjöbring U, Roberts AA, et al. Injury-induced innate immune response in human skin mediated by transactivation of the epidermal growth factor receptor. J Clin Invest 2006; 116:1878 - 85; http://dx.doi.org/10.1172/JCI28422; PMID: 16778986
- Onan D, Allan EH, Quinn JM, Gooi JH, Pompolo S, Sims NA, et al. The chemokine Cxcl1 is a novel target gene of parathyroid hormone (PTH)/PTH-related protein in committed osteoblasts. Endocrinology 2009; 150:2244 - 53; http://dx.doi.org/10.1210/en.2008-1597; PMID: 19147675
- Barman M, Unold D, Shifley K, Amir E, Hung K, Bos N, et al. Enteric salmonellosis disrupts the microbial ecology of the murine gastrointestinal tract. Infect Immun 2008; 76:907 - 15; http://dx.doi.org/10.1128/IAI.01432-07; PMID: 18160481
- Furet JP, Firmesse O, Gourmelon M, Bridonneau C, Tap J, Mondot S, et al. Comparative assessment of human and farm animal faecal microbiota using real-time quantitative PCR. FEMS Microbiol Ecol 2009; 68:351 - 62; http://dx.doi.org/10.1111/j.1574-6941.2009.00671.x; PMID: 19302550