Abstract
Low-copy number plasmids need a segregation mechanism to assort one half of the plasmid copies to each daughter cell during cell division. This can be achieved directly by partitioning plasmid copies through a mechanism reminiscent of eukaryotic mitosis. Briefly, plasmid copies are paired around a centromere-like site, and then separated toward the daughter cells at cell division. Partition mechanisms are used by a majority of well-studied plasmids. They involve two proteins, a DNA-binding protein and a motor protein, besides the centromeric site. However, some plasmids do not encode typical partition systems, so alternative segregation mechanisms must be considered. For instance, chromosome segregation could provide the driving force for plasmid movement, through a “pilot-fish”-like mechanism. In support of this assumption, we recently demonstrated that plasmid R388 segregation, which does not involve a plasmid-encoded motor protein, requires a single plasmid-encoded DNA-binding protein. Besides, the new segregation system becomes essential when the plasmid encodes conjugation machinery, providing a new understanding of how plasmids integrate conjugative transfer with segregation.
Acknowledgments
We thank F. Cornet for helpful discussions. This work was supported by grants BFU2008-00995/BMC from Ministerio de Ciencia e Innovacion (MCINN, Spain), RD06/0008/1012 from Instituto de Salud Carlos III, and 248919/FP7-ICT-2009-4 from the European VII Framework Program. C.G. was the recipient of fellowships from Fondation pour la Recherche Medicale (SPE20080512320) and European Molecular Biology Organization (157-2008).
Figures and Tables
Figure 1 Main types of plasmid segregation systems. The figure shows the genetic organization of the main types of arrangement of plasmid segregation systems, as represented by prototype plasmids P1 (type I), R1 (type II), pBToxis (type III) and R388 (type IV?). Boxes represent genes that encode motor proteins (blue) and DNA-binding adaptor proteins (red). Cis-acting sites are shown as yellow boxes. Dashed curved arrows indicate binding of adaptor proteins to their target sites.
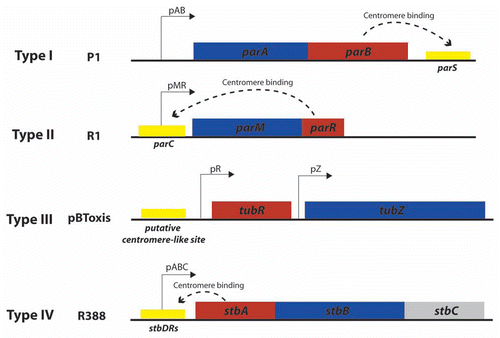
Figure 2 Proposed mechanisms of plasmid segregation. For the sake of clarity, only two newly replicated plasmid copies are represented. When necessary, light red and white regions indicate nucleoid and cytosol spaces, respectively. (A) Pushing mechanism, exemplified by R1 type II par system. Partitioning complexes are formed through specific binding of ParR proteins (red circles) to the centromere-like site parC of newly replicated plasmid molecules, and serve as a nucleation point for ParM-mediated filament formation. Continuous insertion of ParM-ATP motor proteins (blue circles) on the filament ends pushes plasmid molecules apart. Conversion of ParM-ATP to ParM-ADP (open blue circles) leads to destabilization of the filament, thus allowing the entry of another ParM-ATP. At cell division, plasmid molecules localize near opposite cell poles, thus ending in daughter cells.Citation15 (B) Pulling mechanism, as proposed for type I (pB171) and type III (pBtoxis) par systems.Citation16,Citation17 NTP-bound motor proteins (blue circles) bind cooperatively to nucleoid DNA, forming a nucleating core from which filaments form. Subsequently, a growing filament contacts a partition complex formed by the adaptor protein (red circles) bound to the plasmid centromere-like site. Stimulation of NTPase activity of the motor protein by the adaptor protein at the end of the filament leads to conversion to its NDP form (open blue circles) and its release, leaving a new filament end accessible for interactions with the partition complex. The plasmid is thus pulled in the opposite direction to the growth of the filament, and moves around its position, between two other plasmids or between a plasmid and the nucleoid end. (C) Diffusion-ratchet mechanism, as proposed for plasmid P1 type I par system.Citation18 ParB (red circles) loads onto the plasmid at the centromere-like site parS, forming the partition complex. After plasmid replication, partition complexes develop repulsive interactions. ParB stimulates ParA (dark blue circles) ATPase activity, and ParA-ADP molecules (open blue circles) are then excluded from the nucleoid. The motive force for plasmid movement is directed toward regions of high ParA concentration. Movement of the partition complex is thus constrained to one direction because of the low ParA concentration behind it, and at the nucleoid end, it changes direction. ParA-ADP molecules diffuse randomly, exchange ADP for ATP (light blue circles), and then rebind the nucleoid. (D) “Pilot-fish” mechanism, as proposed for plasmid R388 segregation, representing the prototype of a potentially new class of segregation system type IV.Citation21 In contrast to typical par systems, the StbB putative ATPase is not involved in R388 segregation, which does not need a motor protein. We propose that the partition complex, formed by StbA binding to the centromere-like site stbDRs, is used to pair plasmid molecules to the host nucleoid (or other structure associated to the nucleoid). Plasmid segregation is ensured by the host chromosome segregation system.
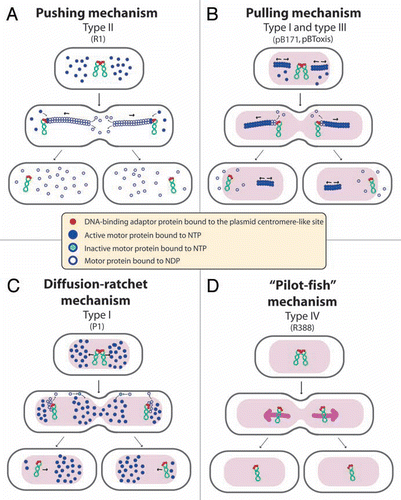
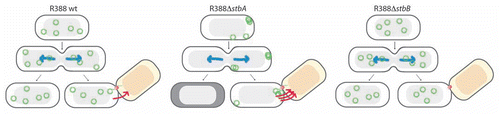
Figure 3 Schematic model to explain the role of the StbAB system.Citation21 Open and shaded regions indicate nucleoid and cytosol spaces, respectively. Blue arrows indicate segregation of the host chromosome, and red arrows represent conjugative transfer to recipient cells (light red nucleoid). Plasmid R388 molecules are evenly distributed both in nucleoid and cytosol areas. At cell division, each daughter cell contains plasmid copies. Localization of plasmid copies at the cell pole is correlated with R388 capacity to undergo conjugative transfer. In contrast, DNA molecules of plasmid R388ΔstbA are exclusively localized in cytosol spaces toward the cell poles and the cell center. This is correlated with plasmid instability, since cells containing all copies in one side of the cell give rise to plasmid-free cells (dark cytoplasm), and with high conjugation frequency, since there are more plasmid copies at the poles. DNA molecules of plasmid R388ΔstbB are distributed in nucleoid but not in cytosol spaces, which correlates with a defect in conjugative transfer.