
Abstract
The extensive and multifaceted traffic between nucleus and cytoplasm is handled by a single type of macromolecular assembly called the nuclear pore complex (NPC). While being readily accessible to ions and metabolites, the NPC imposes stringent selectivity on the passage of proteins and RNA, tightly regulating their traffic between the two major cellular compartments. Here we discuss how shuttling carriers, which mediate the transport of macromolecules through NPCs, cross its permeability barrier. We also discuss the co-existence of receptor-mediated macromolecular transport with the passive diffusion of small molecules in the context of the various models suggested for the permeability barrier of the NPC. Finally, we speculate on how nuclear transport receptors negotiate the dependence of their NPC-permeating abilities on hydrophobic interactions with the necessity of avoiding these promiscuous interactions in the cytoplasm and nucleus.
Figures and Tables
Figure 1 NPC structure and mediated-transport cycles. NPCs fuse the inner and outer membranes of the nuclear envelope (blue), forming aqueous channels that communicate between the nucleus and the cytoplasm. The vertebrate NPC measures about 120 × 90 nm and is made up of ∼30 different proteins, called nucleoporins or Nups, most of which are present in multiples of eight. Associated with the core scaffold of the assembly are eight, ∼50 nm long filaments that protrude towards the cytoplasm and a massive, fish trap-like structure, termed the nuclear basket, which extends about 50 nm into the nucleoplasm. (Left part) Nuclear import. A protein carrying a nuclear localisation signal binds to an import transport receptor in the cytoplasm. Upon reaching the nucleoplasmic face of the pore, binding of RanGTP to the transport receptor frees the latter from FG-repeats in the pore and dissociates the complex. (Right part) Nuclear export. A ternary export complex is formed between a nuclear export receptor, a nuclear export signal-bearing cargo and RanGTP, which typically increases the affinity of export receptors to their cargo. The complex traffics to the cytoplasmic face of the NPC where it is disassembled and the Ran-bound GTP is concomitantly hydrolysed in a process requiring one of two Ran-binding proteins, called RanBP1 and RanBP2 and Ran's GTPase-activating protein RanGAP1. Due to its small size, Ran can, in principle, cross the NPC by passive diffusion. However, to maintain a steep RanGTP gradient across the NE, the transport of GDP-loaded Ran back to the nucleus, where recharging with GTP takes place, is facilitated by a dedicated import receptor called NTF2.
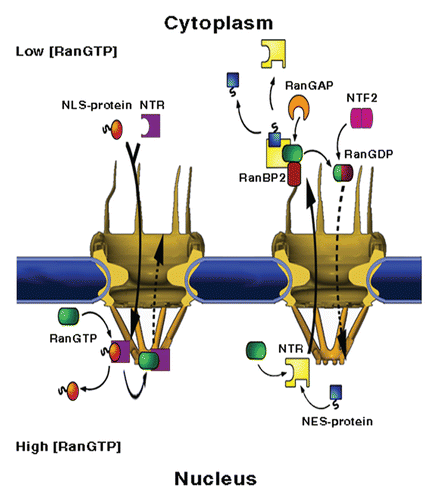
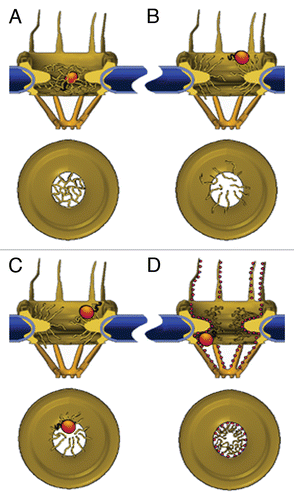
Figure 2 Models of the NPC permeability barrier. (A) Self-interacting FG-repeats form a meshwork within the central pore channel; only molecules that are smaller than the mesh size can go through unassisted. NTRs partition into the meshwork by replacing internal FG-FG contacts with interactions with their hydrophobic patches, virtually ‘dissolving’ into the polymer gel. (B) Unlinked FG-repeats at the edges of the NPCs form polymer brushes that exclude macromolecules through their thermal motion. NTRs permeate this barrier by binding to the FG-repeats, thereby increasing their probability of entry. (C) NTR binding induces collapse of FG-domains, which is reversed by dissociation of the receptor. Transport is afforded by repeated collapse and re-extension events. (D) FG-repeats (shown here as small red spheres) collapse to form a continuous layer extending from the cytoplasmic filaments, through the interior of the central pore channel, to the nuclear filaments. NTRs glide over this surface in a 2D random-walk fashion whereas small molecules pass through an aqueous tube located at the centre of the conductive channel. The space between this tube and the FG-layer in the channel is assumed to be occupied by a loose network of hydrophilic flexible polypeptides, possibly the hydrophilic stretches that connect between the FG-repeats.
Extra View on: Naim B, Zbaida D, Dagan S, Kapon R, Reich Z. Cargo surface hydrophobicity is sufficient to overcome the nuclear pore complex selectivity barrier. EMBO J 2009; 28:2697 - 2705