
Abstract
Inner nuclear membrane (INM) proteins can be important for positioning chromosomes within the nucleus. Little is known about INM proteins in the fission yeast Schizossacharomayces pombe. Telomeres are the most obvious chromosomal sites that are anchored to the nuclear envelope in this organism. A group of proteins that tether telomeres to the spindle-pole body (SPB) during meiotic prophase, such as Bqt1, Bqt2, and Sad1, has been identified previously, but proteins for anchoring telomeres to the nuclear envelope in vegetative cells have not been identified until recently. A recent report demonstrates that Bqt3 and Bqt4 are INM proteins that affect nuclear positioning of telomeres in vegetative cells, and consequently affect the telomere clustering in meiotic prophase. Interestingly, in the absence of Bqt4, telomeres are separated from the nuclear envelope but telomere silencing and telomere length are properly regulated. An important implication of these results is that the functional integrity of telomeres is maintained independently of their connection to the nuclear envelope.
Acknowledgements
We thank Tomoko Kojidani, Kasumi Okamasa, Chihiro Tsutsumi and Miho Yamane for technical assistance. This work was supported by grants from the Japanese Ministry MEXT (Y.C., T.H. and Y.H.).
Figures and Tables
Figure 1 A group of bouquet proteins. (A) Bqt3 and Bqt4 form an attachment surface for telomeres on the nuclear envelope in vegetative cells. (B) Bqt1 and Bqt2 link telomeres to the SUN-KASH protein complex in meiotic prophase.
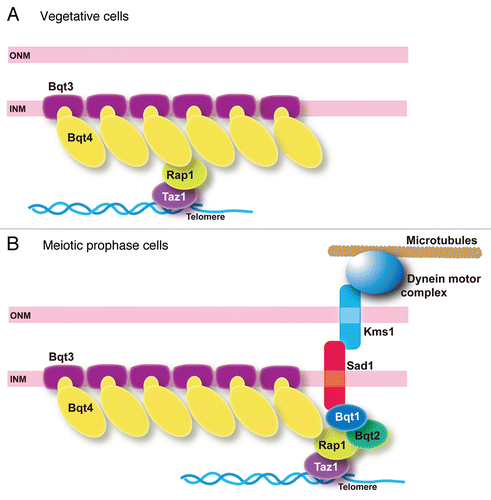
Figure 2 Localization of Bqt4 variants bearing chimeric TM regions. (A) Replacement of the TM sequence of Bqt4 by other TM sequences. The TM of Bqt4 (indicated by the filled box) was replaced by other TM sequences of Sad1 or Lem2 (SPAC18G6.10). Sad1 and Lem2 are INM proteins of S. pombe. Membrane geometry of these proteins is shown in this diagram. The TM of Sad1 and the first TM of Lem2 (indicated by the hatched box) were used to construct the chimeric Bqt4 variants. These TM sequences orient their N-terminus to the nucleoplasm as the TM of Bqt4 does. To construct the chimeric Bqt4 proteins, we replaced the TM of Bqt4 (the C-terminal 19 amino acid residues) with the following sequences: LL WFG IT L FST LLI IT L L (TM of Sad1; 171–188 aa) in Bqt4-Sad1TM, SSY LVH IFM ILL GVV AAI FLA LL (the first TM of Lem2; 316–338 aa) in Bqt4-Lem2TM. (B) Telomere clustering in bqt4Δ cells expressing the GFP-fused Bqt4 variant proteins. Percentages of the nuclei bearing a single cluster of telomeres are shown for each of the variants as indicated. Numbers of cells examined: 124 (GFP-Bqt4-wt), 56 (GFP-Bqt4-dTM), 64 and 85 (GFP-Bqt4-Sad1TM in bqt3+ and bqt3Δ, respectively), 49 and 101 (GFP-Bqt4-Lem2TM in bqt3+ and bqt3Δ respectively). (C) Localization of Bqt4 and its variants bearing chimeric TM fused with GFP in vegetatively growing cells. The bar indicates 10 µm. To express GFP-fused Bqt4 variants, the bqt4 promoter region (1,000 base pair region upstream the coding sequence plus the initial ATG), the sequence encoding GFP-S65T and the coding sequence of the bqt4+ gene, or its variant constructs, followed by the nmt1 terminator sequence were ligated in-frame into an integration vector pYC36;Citation21 the resulting plasmid was integrated into the chromosome at the lys1 gene locus in bqt4Δ cells.
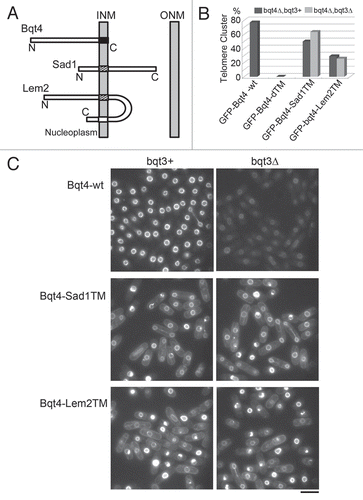
Figure 3 Homologues of Bqt3 and Bqt4 in fungi. Comparison of amino acid sequences of Bqt3 and Bqt4. Identical residues are dark shaded with white letters and conserved residues are light shaded. Species: Sp, Schizossacharomyces pombe; Sj, Schizossacharomyces japonicus; Nc, Neurospora crassa; An, Aspergillus nidulans. Gene ID: Sj bqt3 (SJAG_05226), Sj bqt4 (SJAG_01263), Nc bqt4 (NCU06560) and An bqt4 (AN0162.2).

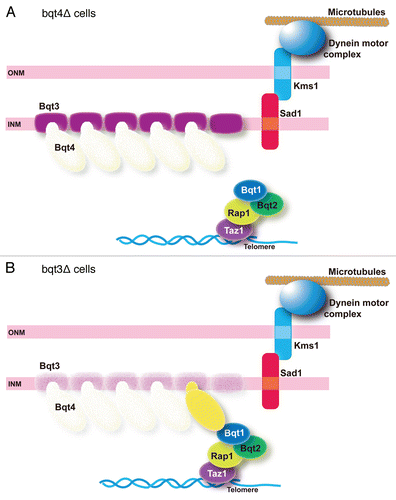
Figure 4 Phenotypes of bouquet mutants. (A) In the absence of Bqt4 (transparent), telomeres are not anchored to the nuclear envelope. However, telomeres may have some chance of contact with Sad1 when telomeres locate near the nuclear envelope through interaction between Bqt1 and Sad1. (B) In the absence of Bqt3 (transparent), degradation of Bqt4 results in basically the same situation as in (A), but residual Bqt4 (yellow) may provide a chance for telomeres to contact with Sad1.
Extra View on: Chikashige Y, Yamane M, Okamasa K, Tsutsumi C, Kojidani T, Sato M, et al. Membrane proteins Bqt3 and −4 anchor telomeres to the nuclear envelope to ensure chromosomal bouquet formation. J Cell Biol 2009; 187:413 - 427