
Abstract
Spatial separation of eukaryotic cells into the nuclear and cytoplasmic compartment permits uncoupling of DNA transcription from translation of mRNAs, and allows cells to modify newly transcribed pre mRNAs extensively. Intronic sequences (introns), which interrupt the coding elements (exons), are excised (“spliced”) from pre mRNAs in the nucleus to yield mature mRNAs. This not only enables alternative splicing as an important source of proteome diversity. Splicing is also an essential process in all eukaryotes and knock-out or knock-down of splicing factors frequently results in defective cell proliferation and cell division. However, higher eukaryotes progress through cell division only after breakdown of the nucleus (“open mitosis”). Open mitosis suppresses basic nuclear functions such as transcription and splicing, but allows separate, mitotic functions of nuclear proteins in cell division. Mitotic defects arising after loss-of-function of splicing proteins therefore could be an indirect consequence of compromised splicing in the closed nucleus of the preceding interphase, or reflect a direct contribution of splicing proteins to open mitosis. Although experiments to directly distinguish between these two alternatives have not been reported, indirect evidence exists for either hypotheses. In this review, we survey published data supporting an indirect function of splicing in open mitosis or arguing for a direct function of spliceosomal proteins in cell division.
Acknowledgements
We thank Beate Faba-Neumann and Thomas Held from the MITOCHECK group at EMBL for continuous help with the MITOCHECK database and the interpretation of phenotypes, and we are grateful to Annette Flotho, Björn Boysen, Bernhard Dobberstein, Georg Stöcklin and Christine Clayton for critically reviewing the manuscript.
Figures and Tables
Figure 1 Dynamic changes of spliceosomal components during the splicing reaction. The spliceosome assembles intermediates during recognition of the splice site (complex A), before and after the first catalytic step (complex B and B*), and for the second catalytic event (complex C). Spliceosomal proteins and subcomplexes are indicated: U1, U2, U4/U6 and U5 snRNPs as well as the nineteen containing complex (NTC) and accessory proteins. The RNAi mediated knock-down of red components leads to defects in mitosis in human cells; green color indicates defects in cytokinesis (see also ).
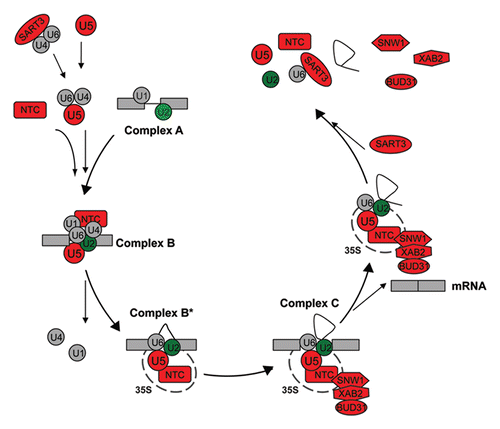
Figure 2 The role of splicing proteins in open mitosis. During Interphase (top) the nuclear envelope ensures spatial separation of nuclear and cytoplasmic processes and NPCs (Nuclear Pore Complex) mediate exchange of molecules between the cytoplasm and the nuclear compartment. Special areas in the nucleus harbor many splicing proteins (“splicing speckles”, top, gray gradient). With the onset of mitosis (arrows) phosphorylation of lamins and NPC proteins triggers the disassembly of the nucleus. Some components of the NPC directly contribute to open mitosis; their knock-down results in cell division defects (bottom, right). These are relocalized to the mitotic spindle or kinetochores (bottom). In contrast, splicing factors are evenly distributed in the open mitotic cytoplasm (bottom, gray gradient). However, their knock-down also leads to defective mitoses (bottom, center).
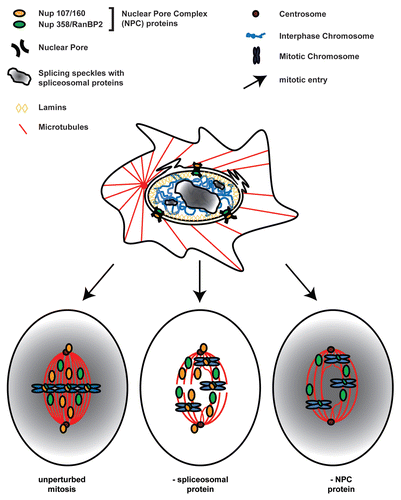
Figure 3 The Molecular basis for mitotic defects after downregulation of splicing proteins. (A) The knock-down of spliceosomal proteins (gray gradient) causes splicing defects, which are causative for mitotic defects. Left: unperturbed mitosis; all mitotic gene products, like e.g., the cell cycle regulator CDC25, are properly spliced and active in mitosis. Middle: Impaired splicing and subsequent loss-of-function of a single gene product, exemplified by CDC25, causes mitotic defects. Right: Impaired splicing leads to the accumulation of improperly processed bulk mRNAs (yellow), which in turn compromise chromatin organization and chromatin condensation in mitosis, and interfere with mitotic progression. (B) Mitotic defects after knock-down of splicing proteins (red) arise as a consequence of direct functions of splicing proteins in open mitosis. Left: unperturbed mitosis; splicing proteins work in the interphase nucleus (red color) and independently in mitosis (yellow color). Right: Their knock-down leads to mitotic defects directly.
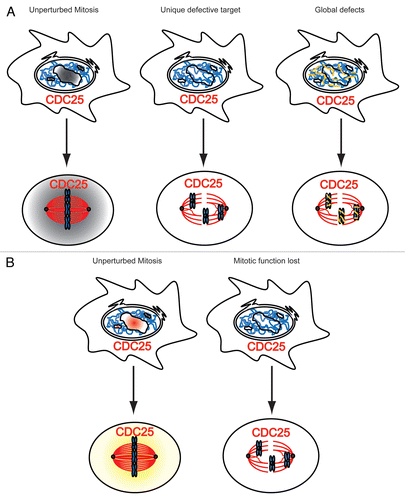
Table 1 Knock-down phenotypes of spliceosomal components in human cells